


Reddish Egret (Egretta rufescens) in the Lower Florida Keys
Abstract
The Reddish Egret (Egretta rufescens) is a species that is greatly dependent on shallow tidal flats ideal for its active foraging behavior. It is unknown why some seemingly identical islands are chosen for nesting over others. The goal of this study is to determine if there is a difference in amount of available foraging habitat between mangrove keys in the Lower Florida Keys that do and do not have Reddish Egrets nesting on them within a 5 km, 10 km, and 15 km radius using a geographic information system (GIS). Islands with nesting Reddish Egrets had a greater amount of foraging habitat within a 5 km radius (p = 0.027) while at a radius of 10 km and 15 km there was no difference (p = 0.29, p = 0.32 respectively) between islands containing nesting Reddish Egrets and those without nests. Our results suggest that at short distances of <5 km, the amount of foraging habitat is important to adult Reddish Egrets choosing an island/nesting location. Protection of foraging habitat in spatial proximity to nesting colonies is essential to protecting and managing this species in the future.
Key words: Ardeidae; colony; geographic information system.
Introduction
The availability of adequate resources is necessary to sustain populations of animals. Biologists and managers are often concerned with the amount of resource available and the differential use of resources. Availability is the quantity of a resource that is accessible by the animal or population during a given period of time (Manly et al. 2002). Assessing resource usage and availability is important especially for threatened or endangered species as understanding the resources that are important to the animal can provide useful information in the management and protection of that species. The availability of foraging habitat and resources may be largely influential in breeding site location. For those organisms with prime foraging habitat separated from breeding habitat, the availability of foraging habitat may strongly dictate the use of specific breeding habitat.
The Reddish Egret (Egretta rufescens) is an excellent example of an organism which requires two distinct, separate habitats. With its highly restricted geographic distribution and narrow habitat requirements, the Reddish Egret has an estimated global population of 5,000-7,000 breeding pairs (Paul 1991, Green 2006). Currently in Florida, the Reddish Egret is considered a “species of special concern” mainly due to plume hunting around the end of the 19th century and the beginning of the 20th century that nearly extirpated the population. Current threats including habitat degradation and loss, presence of predators and human disturbance have slowed the recovery of the population (Lowther and Paul 2002). Current estimates for Florida put the population at approximately 300 breeding pairs (Green 2006). Reddish Egrets are restricted by the availability of foraging habitat comprised of broad, saline coastal flats, ideal for its active foraging behavior (Paul 1991). The Reddish Egret typically forages in depths less than 20 cm (Farmer 1991) and rarely up to 25 cm (Lowther and Paul 2002). The diet is usually made up of small fish that occur at these shallow depths as shown by nestling regurgitations in the Laguna Madre that were comprised of mostly sheepshead minnow (Cyprinodon variegatus; 83%), longnose killifish (Fundulus similis; 9%), pinfish (Lagodon rhomboides; 5%); also striped mullet (Mugil cephalus) and ladyfish (Elops saurus; McMurry 1971, Simersky 1971, Holderby and Green unpubl. data).
The Reddish Egret is a year round resident along the coastlines of Texas, Florida, the Pacific and Gulf Coasts of Mexico, and the Bahamas (Cook 1913, Lowther and Paul 2002). Nesting takes place on islands to presumably avoid terrestrial predators (Paul 1991). In Florida nesting takes place on mangrove keys consisting primarily of red mangrove (Rhizophora mangle) with nests typically placed at least a few meters up from the ground or water (Lowther and Paul 2002). There are many gaps in what is known about Reddish Egret biology including why particular islands or nesting locations are chosen over others that are seemingly quite similar. We hypothesize that the availability of foraging habitat is influential in the selection of mangrove key islands for nesting/ breeding in the Lower Florida Keys (LFK) and that the amount of foraging habitat closer to an island is more influential than foraging habitat further away in the decision where to nest. Assessing how the availability of foraging habitat influences nest site selection will contribute to the understanding of this coastal wading bird and aid management objectives for these threatened birds and their coastal habitat in the future.
Methods
Two surveys were conducted in the Lower Florida Keys during the second week of March and the last week of May in 2007. Surveys were conducted on islands historically occupied by Reddish Egrets (Powell et al. 1989); islands were surveyed by encircling each island and brief forays on to the island in search of Reddish Egrets. Islands were identified as a nesting island if we detected the presence of active nests (e.g., eggs, nestlings) or if hatchling year Reddish Egrets were seen on the island. If no nestlings or hatchling year birds were detected then the island was assumed to be absence of Reddish Egrets. Hatchling year birds were identified as those that had recently fledged and had the ability to fly but had not yet left the nesting island. They were also usually characterized by smaller size and dull plumage. The islands surveyed include Budd Key, Marjoe Key, Crawl Key, Niles Channel Key, Cupon Bight Key, Sawyer Key, Torch Key, Picnic Key, Happy Jack Key, Galdin Key, and Little Pine Key (Powell et al. 1989). It is important to note that for the purposes of this study it is assumed that the influence of predators and nest site quality was the same on every island and that the quality of available foraging habitat was similar.
To address this hypothesis, we used a geographic information system (GIS) to calculate the area of foraging habitat within a 5, 10 and 15 km radius of the surveyed islands. Using ArcGIS (ESRI, Redlands, California, USA), we imported geographic layers and bathymetry data for the Lower Florida Keys from the Marine Resources Geographic Information Systems by Florida Fish and Wildlife Conservation Commission. The bathymetry data shows contours of the depths that occur in the LFK including contours that indicated Mean Low Water, contour depth < 1m. We deemed this contour suitable for representing foraging area as the land is only exposed at the average of the low tides and is thus covered with water the rest of the time and presumably available foraging habitat. Using this bathymetry data, a new layer was created to represent available foraging habitat. We then overlaid concentric 5, 10, and 15 km buffers around each colony and used this layer as available foraging habitat within each concentric buffer. After creating the layer resulting from the intersection of the buffer and the foraging habitat layer, the total available foraging area within each buffer was calculated for each island. To test the null hypotheses that the amount of available foraging area was not different between islands with and without nesting Reddish Egrets, we performed a t-test (program R, The R Foundation for Statistical Computing, 2006) between occupied and unoccupied islands at distances of 5, 10, and 15 km.
Results
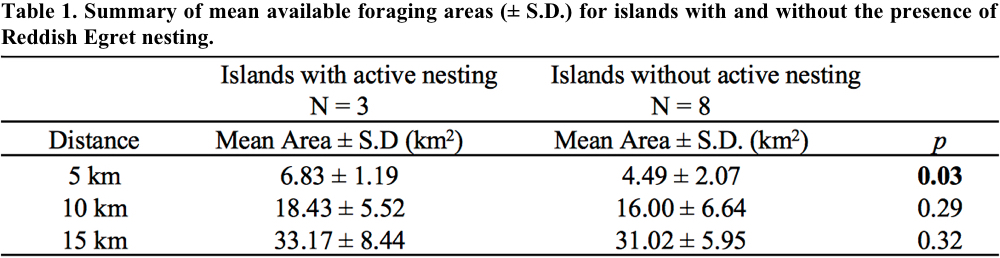
We surveyed all historically known nesting islands for Reddish Egrets and documented nesting on only Budd Key, Picnic Key, and Happy Jack Key; all three islands exhibited signs of Reddish Egret nesting by evidence of nestlings or hatchling year birds. We failed to detect any signs of nesting on the remaining 8 islands. The mean available foraging area within 5 km for islands with Reddish Egrets was greater than the mean available foraging area within 5 km for islands without Reddish Egrets (Table 1). For distances of 10 and 15 km, we detected no significant difference between available foraging area for islands with and without nesting Reddish Egrets.
Discussion
Within a 5 km radius of mangrove islands in the LFK, amount of available foraging habitat is apparently an important factor for Reddish Egrets in selecting suitable islands for nesting; available foraging habitat outside of 5 km radius did not appear to influence island selection. Shorter distances to available foraging habitat is advantageous to Reddish Egrets as it is more energetically efficient and allows more time for parental care at the nest. Nestlings on islands with more available foraging habitat in closer spatial proximity presumably means shorter time interval between feeding of young as time of flight is reduced for adults traveling to foraging areas. One reason why available foraging area may not have been significantly different at 10 and 15 km radii is because foraging areas at this distance from the nesting island tended to overlap with the foraging area for other islands resulting in the same foraging area being used my multiple islands. This fact may violate the assumption of statistical independence. Future studies should include more sampling sites (e.g., Florida Bay, Upper Florida Keys) so that islands can be chosen that have spatial independence and do not share foraging habitat within a given radius (15 km). Historically, Reddish Egret nesting in the Florida Keys and Florida Bay has two main pulses from November to February and from February to May and there is the potential that we missed birds by only surveying twice during the latter pulse (Lowther and Paul 2002). Also, conducting this study in other regions including Laguna Madre of Texas and Mexico may yield different results as nesting colonies are typically less densely concentrated than islands in the LFK.
We did not examine other factors potentially related to colony site selection including factors such as prey availability, presence of predators and human disturbance. Future studies may benefit from analyzing more variables such as prey composition and density, predator composition and density, presence of other nesting waterbirds and the frequency and extent of human disturbance. Also, all of these islands are subjected to storms and hurricanes which could result in differential habitat quality between islands from the effects of these weather events. Regardless, our coarse-scale analysis does demonstrate the importance of available foraging habitat within close spatial proximity to nesting sites and provides an impetus for conservation of not only nesting islands, but the surrounding available foraging areas.
Acknowledgements
We thank J. Lorenz, T. Wilmers, S. Melvin, the Florida Fish and Wildlife Commission, the Florida Keys National Wildlife Refuge System and the U.S. Fish and Wildlife Service. Funding for this study was provided by U.S. Fish and Wildlife Service, Region 4 through a grant to MCG.
Literature Cited
Cook, W. W. 1913. USDA Biological Survey, Bulletin No. 45. 70 pp.
Farmer, M. 1991. Reddish egrets of the lower Laguna Madre, Texas. U.S. Fish and Wildlife Service, Lower Rio Grande National Wildlife Refuge, Texas, U.S.A.
Green, M. C. 2006. Status Report and survey recommendations on the Reddish Egret (Egretta rufescens). U.S. Fish and Wildlife Service, Georgia, U.S.A.
Lowther, P. E. and R. T. Paul. 2002. Reddish Egret (Egretta rufescens), The Birds of North America Online (A. Poole, Ed.). Ithaca: Cornell Lab of Ornithology; Retrieved from the Birds of North America [online].
Manly, F. J., McDonald, L. L., Thomas, D. L. and McDonald, T. L. 2002. Resource selection by animals. Kluwer Academic Publishers, New York, U.S.A.
McMurry, S. L. 1971. Nesting and development of the Reddish Egret (Dichromanassa rufescens Gmelin) on a spoil bank chain in the Laguna Madre. M.S. Thesis, Texas A&M University, Texas, U.S.A.
Paul, R. T. 1991. Status Report - Egretta rufescens (Gmelin) Reddish Egret. U.S. Fish and Wildlife Service, Texas, U.S.A.
Powell, G. V. N., R. D. Bjork, J. C. Ogden, R. T. Paul, A. H. Powell and W. B. Robertson, Jr. 1989. Population trends in some Florida Bay wading birds. Wilson Bulletin 101:436-457.
Simersky, B. 1971. Competition and nesting success of four species of herons on four spoil islands in the Laguna Madre. M.S. Thesis, Texas A&M University, Texas, U.S.A.