


Changes of the nesting sites of Grey Herons (Ardea cinerea) in Hokkaido, northern Japan #
Abstract
The objective of this study was to identify the habitat of all known nesting colonies of the Grey Heron (Ardea cinerea) in Hokkaido, northern Japan, during the period 1960 to 2016. During the study period, 153 colonies were confirmed. All herons nested in trees contiguous with the surrounding land until the early 1990s. After the mid-1990s, however, nine colonies were confirmed in atypical locations: flooded trees, buoys in waters and offshore islets. These locations were considered to be selected to defend their nests against terrestrial predators by being surrounded by water. I suggest, therefore, that securing environments that cannot be physically approached by terrestrial predators should be a priority to conserve the habitat of Grey Herons in Hokkaido.
Key words: Ardea cinerea; Brown Bear; Common Raccoon; Grey Heron; Hokkaido; nesting site; predation; terrestrial predator.
# This paper was presented at the 1st Herons of the World Symposium at the 40th Anniversary Meeting of the Waterbird Society at New Bern, North Carolina, USA, 21-23 September 2016. Other papers from that Symposium will appear in future issues of the Journal of Heron Biology and Conservation, and Waterbirds.
Introduction
Grey Herons (Ardea cinerea) are known to nest in various habitats other than trees, for example, cliff ledges, reed beds and on the ground on islands (Lowe 1954, von Westernhagen 1970, Litvinenko 1982, Roalkvam 1994, Thomas and Hafner 2000). Variation in nesting substrate is similarly seen in Great Blue Herons (Ardea herodias) (Oberholser 1912, Gabrielson and Jewett 1940, Hancock and Elliot 1978, Burger 1979, Sandilands 2010).
Two explanations have been proposed for these atypical nesting sites. One is a lack of trees. Thomas and Hafner (2000) described the change of nesting habitats of Grey Herons from trees to reed beds after severe logging. Henny and Kurtz (1978) also observed that Great Blue Herons nested on channel markers and on the ground on a river islet after the loss of nesting trees.
Another explanation is terrestrial predators. Hafner (1997) noted that heron nests must either have a surrounding security zone, or be in high trees to protect the nests against predators. However, nests in trees are often vulnerable to predators that can climb trees. In such cases, herons may nest where surrounded by a security zone such as water.
In Hokkaido, Grey Herons have recently begun to nest in atypical locations. Although some of the nests were still in trees, all of the colonies were surrounded by water. I propose that the herons chose such environments to avoid terrestrial predators because there was no shortage of trees around the colonies.
In this paper, I describe some atypical types of nesting sites of the Grey Heron in Hokkaido, and discuss the effect of terrestrial predators on nest site selection of herons.
Methods
For data on colonies confirmed by 2005, I referred to the following reports: Matsunaga et al. (2000) and Matsunaga (2005), in which 122 colonies were confirmed. Subsequently, I continued to collect information from ornithologists, birders and museums, and acquired information on another 31 colonies. I visited 25 of those 31 sites to count nests, record the environmental characteristics of the nesting site and obtain information on the year of initial colony formation from local residents. In the case of colonies surrounded by water, I calculated the distance between a colony and the nearest shore using Google satellite map. Because this study is still ongoing, I have not yet conducted field surveys for the remaining six colonies. However, I confirmed that those colonies are not surrounded by water.
Study Area
Field surveys were conducted throughout Hokkaido, Japan.
Results
Between 1960 and 2016, 153 Grey Heron colonies were confirmed in Hokkaido (Fig. 1). Only Grey Herons nested in 147 of the colonies; however, at six sites, Grey Herons nested with other colonial species: with Black-crowned Night-Herons (Nycticorax nycticorax) at three sites, with Great Cormorants (Phalacrocorax carbo) at two sites and with a pair of Great Egrets (Ardea alba) at one site.
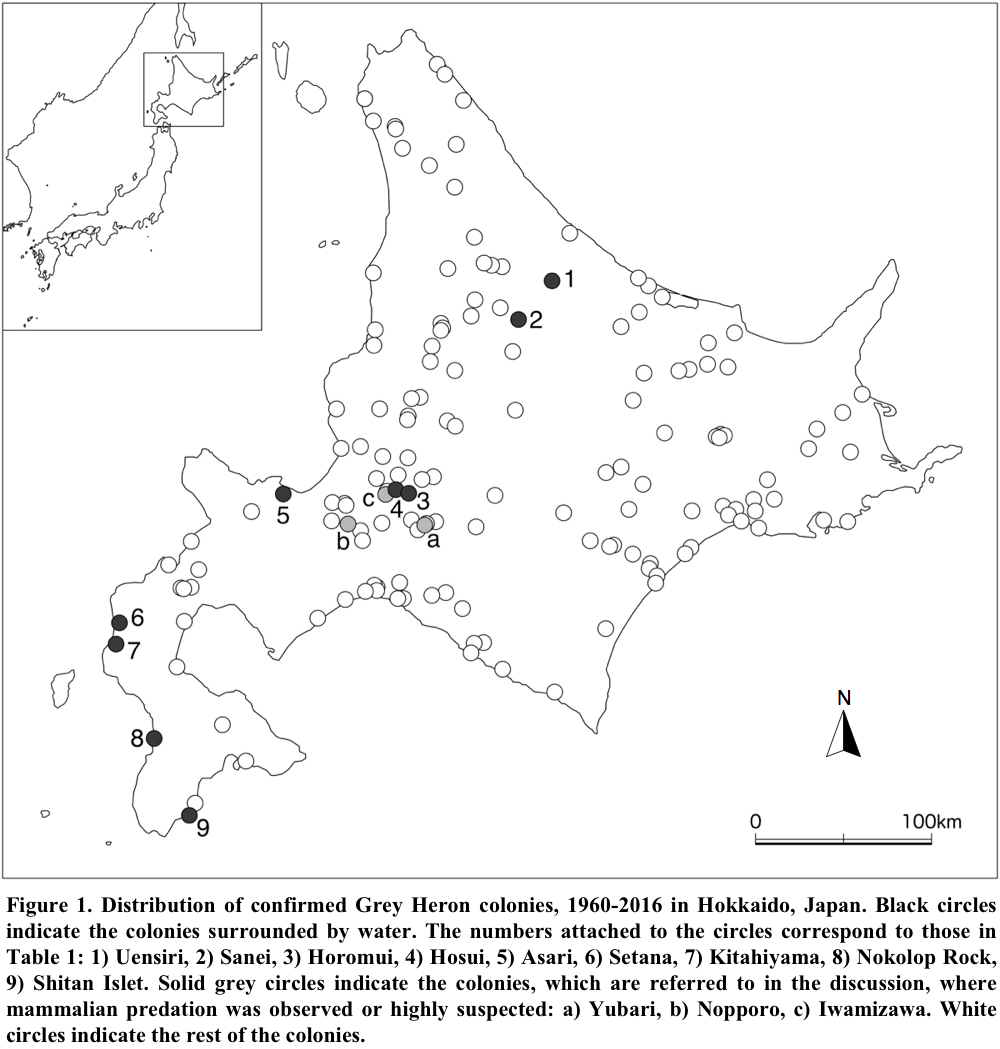
In all 58 colonies located by 1990, herons nested in woods that were not surrounded by water. Since the mid-1990s, however, nine colonies out of 95 colonies have been confirmed in areas surrounded by water (Table 1).

Seven of those nine colonies were located in reservoirs and a small pond. In three of them, herons nested on willow bushes (Fig. 2), which became submerged, except for the top of the bushes, during the breeding season. The nests in two other colonies were placed in dead trees half submerged throughout the year. In two additional colonies in reservoirs, herons nested on buoys (Fig. 3), although, to be precise, a pair of herons in one colony placed their nest on an iron stage built on some buoys. The two remaining colonies were on two offshore islets. Both islets were small and uninhabited. One of them, Nokolop Rock (approximately 65 m long, 33 m wide and 15 m high) is a rocky islet located about 300 m from the mainland. It had no trees but small patches of low herbaceous plants. All the nests observed were on the ground. Some nests were placed on a ridge and others were hung off the side of a cliff (Fig. 4). The other offshore islet, Shitan Islet (approximately 62 m long, 21 m wide and 30 m high), was about 80 m offshore. It had only two dead trees on the top and its steep slope was partly covered by Giant Knotweeds (Fallopia sachalinensis) and Poaceae sp. Three nests were observed in the trees and three others were on the ground in low vegetation.



Discussion
Undoubtedly the lack of trees will force herons to nest in atypical sites. However, this does not apply in Hokkaido, because all of the colonies examined have a sufficient amount of woods around them. Thus terrestrial predators are highly suspected of causing the herons to move to these atypical nesting sites. Although this study did not initially focus on predators, I obtained reliable information about predation and I also witnessed an incident described below.
Information was given to me by a local resident living several hundred meters away from the Yubari colony (Fig. 1), where I previously confirmed 68 nests in November 1993 (Matsunaga 2005). According to his observation, Brown Bears (Ursus arctus) often appeared in the colony in 1994 and 1995. He said that bears climbed the nest trees, repeatedly caught nearly fledged young and, on at least one occasion, knocked the nest down. Consequently, herons did not return to the colony in the next breeding season.
I do not know of any other reports regarding Grey Heron predation by Brown Bears, but a Russian colony was supposedly attacked by Brown Bears on the basis of claw marks of bears on nesting trees (Pronkevich et al. 2012). In the case of the Great Blue Heron, the American Black Bear (Ursus americanus) is noted as a predator (Foss 1980, Gebauer and Moul 2001).
Common Raccoons (Procyon lotor) could be another terrestrial predator in Hokkaido. At the end of April 1997, a Nopporo colony, where 266 pairs of Grey Herons nested in 1990, was suddenly abandoned (Matsunaga 2005). Ikeda (1999) suspected that the colony had been subjected to raccoon depredation because there were many claw marks on nesting trees. In addition, around noon on 30 May 2012, I observed in the Iwamizawa colony, where at least 65 pairs nested in 2004 (Matsunaga 2005), that a raccoon climbed nesting trees and stayed at some nests for more than 30 minutes. I could not see the inside of the nest but infer that the raccoon was eating chicks because it continued tearing up something in the nest. Several days after the incident, I wrapped the nesting trees with metal flashing sheets and, consequently, the herons have continued to nest so far.
I could not find any other reports of raccoon predation on Grey Herons, but Lopinot (1951) and von Duyke (2006) observed that raccoons ate nest contents of Great Blue Herons in the United States where raccoons are native.
In Hokkaido, Brown Bears are estimated to have increased about 1.8 times between 1990 and 2012 (Hokkaido Prefectural Government 2015). On the other hand, the Common Raccoon, which is an introduced species from North America, became widely established in the wild in Hokkaido since the late 1980s (Ikeda 1999), and thereafter their habitat range had expanded over most of Hokkaido by 2016 (Hokkaido Prefectural Government 2016). In addition, Grey Herons also were estimated to have increased about 4.5 times between 1960 and 2005 in Hokkaido (Matsunaga 2005). Thus, it is likely that herons would have encountered bears and raccoons more frequently recently than before, and some herons would, accordingly, have moved to the places where ground predators cannot access their nests.
I could not judge which predators were responsible for the abandonment of each colony. However, it seems more likely that bears attacked at least three of the nine colonies indicated in the results, because raccoons were not confirmed in those regions at the time that the colonies were found.
One should note, the alternative nesting sites are not necessarily surrounded by water. For example, herons in Yubari, after the depredation by bears, founded their new colony close to a house. Some nests were only several meters from it and one nest even had a dog house directly underneath. I presume herons may have used the dogs as deterrents to potential predators. Herons around the Nopporo colony also nested in places close to a residential area after the presumed attack by raccoons. Three new colonies were located within 20 km of the Nopporo colony just after its abandonment, though these new colonies may have been independent of the Nopporo colony. Although the Nopporo colony was located in a large woods, two of the new colonies were situated in small groves surrounded by residential area, and one of them was in a small grove between a water and residential area. These could be cases where human presence (instead of water) works as a barrier against predators.
Because there are not many isolated environments surrounded by water in Hokkaido, more herons may be forced to nest in residential areas, which may cause further problems between the herons and the residents. Thus, it should be a priority to secure isolated environments surrounded by water especially in the region where such habitats are scarce like Hokkaido.
Acknowledgements
This study would not have been possible without the support and cooperation of many people who provided valuable information about colony sites. It is difficult to name all of them, but I would especially like to thank: Hiroshi Echigo, Jun Matsuyama, Hidefumi Mizuma, Hidetoshi Nagano, Atsushi Okuda, Kazuhiro Oodate, Genshi Sakakibara, Mitsuaki Sugita, Junichi Terashima, Toru Tomikawa and Chikashi Utsumi.
Literature Cited
Burger, J. 1979. Resource partitioning: nest site selection in mixed species colonies of herons, egrets and ibises. American Midland Naturalist 101: 191-210.
Foss, E. 1980. A Black Bear in a Great Blue Heron colony. Murrelet 61: 113.
Gabrielson, I. N. and S. G. Jewett. 1940. Birds of Oregon. Oregon State College, Corvallis, Oregon, U.S.A.
Gebauer, M. B. and I. E. Moul. 2001. Status of the Great Blue Heron in British Columbia. Wildlife Working Report No. WR-102. Ministry of Environment, Lands and Parks Wildlife Branch, Victoria, British Columbia, Canada.
Hafner, H. 1997. Ecology of wading birds. Colonial Waterbirds 20: 115-120.
Hancock, J. and H. Elliot. 1978. The herons of the world. Harper and Row, New York, New York, U.S.A.
Henny, C. J. and J. E. Kurtz. 1978. Great Blue Herons respond to nesting habitat loss. Wildlife Society Bulletin 6: 35-37.
Hokkaido Prefectural Government. 2015. Higuma seisokusu no suitei nitsuite. [online] Accessed 31 January 2017.
Hokkaido Prefectural Government. 2016. Araiguma no seisoku shichosonsu to hokakusu to no suii. [online] Accessed 31 January 2017.
Ikeda, T. 1999. Progress of naturalization of raccoons and related problems in Hokkaido. The Annual Report on Cultural Science, Faculty of Letters, Hokkaido University 47: 149-175.
Litvinenko, N. 1982. Nesting of Grey Heron (Ardea cinerea L.) on sea islands of South Primorye. Journal of Yamashina Institute of Ornithology 14: 220-231.
Lopinot, A. C. 1951. Raccoon predation on the Great Blue Heron, Ardea herodias. Auk 68: 235.
Lowe, F. A. 1954. The heron. Collins, London, U.K.
Matsunaga, K. 2005. Status report of the Grey Heron in Hokkaido. [online] Accessed 31 January 2017.
Matsunaga, K., A. Matsuda and H. Fukuda. 2000. Changing trends in distribution and status of Grey Heron colonies in Hokkaido, Japan, 1960-1999. Japanese Journal of Ornithology 49: 9-16.
Oberholser, H. C. 1912. A revision of the forms of the Great Blue Heron (Ardea herodias Linnæus). U.S. Government Printing Office, Washington D.C., U.S.A.
Pronkevich, V. V., A. L. Antonov, A. J. Oleinikov and K. N. Tkachenko. 2012. A new colony of Grey Heron Ardea cinerea Linnaeus, 1758 in the middle Amur River. Amurian Zoological Journal 4: 401-402.
Roalkvam, R. 1994. Gråhegre Ardea cinerea. Pp. 52-53 in Norsk Fugleatlas: Hekkefuglenes Utbredelse og Bestandsstatus i Norge (J. O. Gjershaug, ed.). Norsk Ornitologisk Forening, Trondheim, Norway.
Sandilands, A. 2010. Birds of Ontario: habitat requirements, limiting factors, and status. Nonpasserines: waterfowl through cranes. UBC Press, Vancouver, British Columbia, Canada.
Thomas, F. and H. Hafner. 2000. Breeding habitat expansion in the Grey Heron (Ardea cinerea). Acta Oecologica 21: 91-95.
von Duyke, A. 2006. Monitoring, research, and management activities at the Peltier Island heronry during 2004 & 2005 field seasons. State Wildlife Grants Program, Division of Ecological Services Minnesota Department of Natural Resources. [online] Accessed 31 January 2017.
von Westernhagen, W. 1970. Über die Brutvögel der Banc d’Arguin (Mauretanien). Journal of Ornithology 111: 206-226.