


A new heron colony in Yunnan, China, survives amid the COVID-19 pandemic
Abstract
Urban landscapes with large vegetative structure and water features are a preferred habitat type for colonial water birds including the herons (Ardeidae). However, the aggregation of these birds and their colony growth are often considered a nuisance to the local environment. The resulting human-heron conflicts may ultimately lead to abandonment of the nest-sites and colony displacement. It is therefore important to track the developmental history of heron colonies in relation to the humans and to identify key habitats for preserving and better-managing the urban heron populations. In 2011, two nests of Black-crowned Night Herons (Nycticorax nycticorax) were found in trees of Kunming Zoo, Yunnan Province, China. By 2021, the colony had grown to 418 nests with Little Egret (Egretta garzetta) and Black-crowned Night Heron as the dominant species and with Intermediate Egret (Ardea intermedia) and Great Egret (Ardea alba) also joining the annual breeding season. Chinese Pond Heron (Ardeola bacchus), Cattle Egret (Bubulcus ibis) and Grey Heron (Ardea cinerea) were also sighted, but no or very little nesting by those species was observed. Colony relocation occurred several times, but mainly from trees on the mainland to two artificial pond islands, then from the islands to a mainland grove. At the mainland habitat, nesting by the herons conflicted with 1-2 annual commercial markets and with pigeon-feeding by tourists (in 2021). The former caused negative hygienic concerns as well as increased the management workload. It was thought that continued colony expansion and the increasing heron-human conflicts would eventually lead to a point where colony survival could be jeopardized. In 2020, instead, due to the global outbreak of the COVID-19 and subsequent temporary closure of the park with restriction on public aggregations, growth of the colony seemed to have accelerated. In 2021, with the resumption of the park functions following local Covid case-reductions, enhanced anti-heron actions were imposed in response to an enlarged heron colony, which then might negatively impact the heron breeding success. In this article, we present the developmental history of the heron colony in Kunming Zoo using a timeline, and conclude that colony fate can be largely subject to a series of stochastic events due to changing relationships with the humans. The article serves as case study and reference to a number of other, currently under-studied colonies in urban China, where investigations of human-wildlife relationships are recommended for the future conservation of urban wildlife.
Key words: China; COVID-19; developmental history; human-heron conflicts; Kunming Zoo; urban heronry.
Introduction
Heron populations can serve as an ecological indicator of an improved, or deteriorated, urban environment (Kushlan 1993, Rahman and Ismail 2018) with aesthetic values (Hall 1985). However, their increase and aggregation are frequently treated as a nuisance putting local environmental health at risk (Nisbet 2000). Individual colonies may persist for decades if left unperturbed in/around the same site with the potential to grow in size following establishment (Mashiko and Toquenaga 2018).
In China, where colonies of one or more heron species are sometimes found in the highly-urbanized part of a city (e.g. urban parks), conflicts — resulting from an overlap of wildlife inhabitation and human land-uses — have become an increasing management challenge (Yan et al. 2006). Since urban habitat patches (Angold et al. 2006) are usually of limited space for colony development, colony expansion within the available sites will often further the damage to the nest-supportive vegetation and may also hinder the vegetation regeneration processes (Fedriani et al. 2017).
In response to inhabitation by the nuisance heron colonies, trimming and harassments of nest trees and birds are sometimes adopted which introduce disturbances and may cause mortalities of the eggs and young in active tree-nests; such actions could eventually lead to colony relocation (Shen et al. 2018). In other cases, direct interferences made to the breeding pairs and nestlings would draw criticism as well as lead to legal consequences, since it is illegal to capture or kill herons. However, there are many cases where colony relocation, due to anthropogenic causes, have escaped public records (Wong et al. 2004). It is thus suggested that urban heron colonies be studied both in terms of geographic distribution, associated with land-use types (Carrasco et al. 2017), and of temporal dynamics in one or more focal groups (Dusi and Dusi 1987), in order to understand habitat selection and the relationship between colony growth and human responses.
The global outbreak of COVID-19, in the spring of 2020, and the subsequent lock-down (as well as re-opening) of the public facilities resulted in a significant impact on traditional human-wildlife relationships on a global scale, for example, research on the consequences of the “great human pause” (Rutz et al. 2020). In this article, we present the first case study of an urban park heron colony with a traceable developmental history (2011-2021), including the COVID-19 lock-down era in Kunming Zoo, Yunnan Province, China. As gaps can currently be found in the ecology of urban wildlife in the majority of mainland Chinese provinces, we aim to raise future research interests in this field, which potentially could improve the management of the urban wildlife and their key habitats. This colony was comprised mainly of nesting Little Egrets (Egretta garzetta), Black-crowned Night Herons (Nycticorax nycticorax, henceforth, Night Herons) and smaller numbers of nesting, or visiting, Intermediate Egrets (Ardea intermedia), Great Egrets (Ardea alba), Cattle Egrets (Bubulcus ibis), Grey Herons (Ardea cinerea) and Chinese Pond Herons (Ardeola bacchus).
Study Area
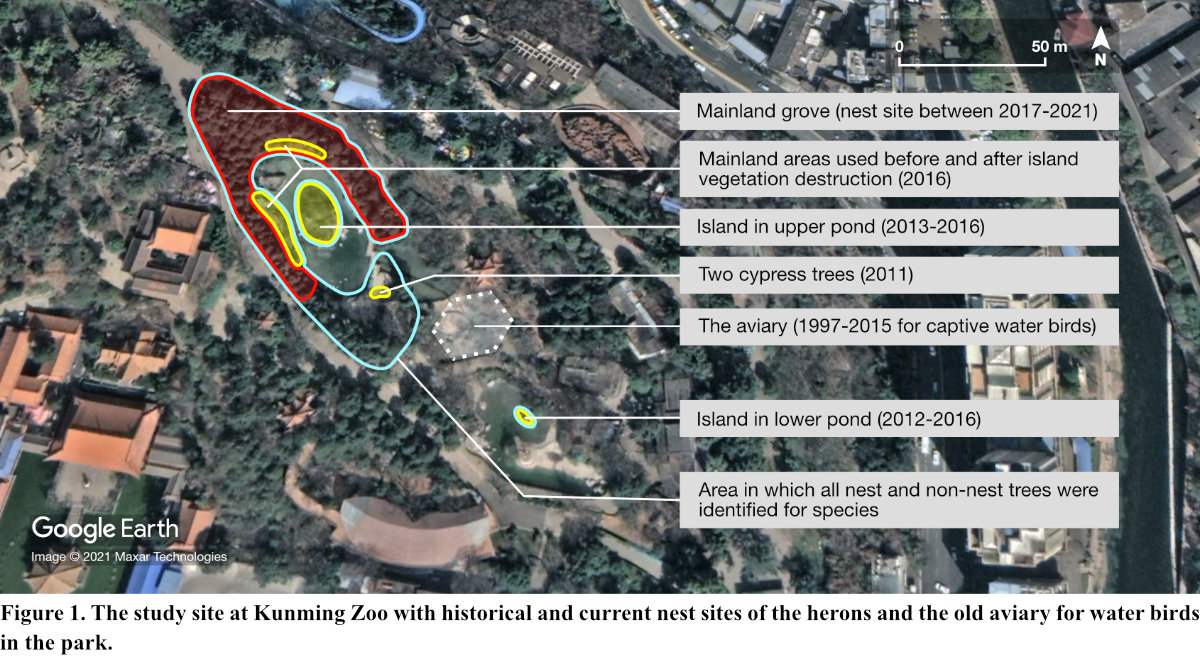
Kunming Zoo (25° 03′ 12″ N, 102° 42′ 45″ E, elevation: 1,910 m) is situated at Mt. Luofeng — an urban highland characterized by karst terrains (Zhang 2001) — in downtown Kunming, Yunnan Province, southwest China. Both entertainment facilities (e.g., theme parks, cafeterias, booths) and zoo enclosures are present. The vegetation cover of the park is largely a mixture of plantation and remnant forests of relatively continuous canopies, with woody strips, bamboos, shrubs and grasses within the 23 ha of the park territory.
The historical and current nest sites for herons were located in the Water Bird Sector of the park and were composed of two water ponds (an upper and a lower pond) and the adjacent mainland surrounding the upper pond, including a small cypress grove to the north (Fig. 1). Each pond had an artificial island offering vegetative support to the nesting herons. At the nest sites on the islands and some waterside areas in the adjacent mainland, which were used until 2016, at least seven different tree species were identified in 2020. They included: Chinese Cypresses (Cupressus duclouxiana), Weeping Willows (Salix babylonica), Broad-leaved Privets (Ligustrum lucidum), Waxy Leave Privets (L. quihoui), Paper Mulberries (Broussonetia papyrifera), Hall Crabapples (Malus halliana), Himalayan Cherries (Cerasus cerasoides) and Crape Myrtles (Lagerstroemia indica). All of the trees besides the Crape Myrtles (often short and probably not suitable for nesting in) were observed or assumed to have once served as nest trees before colony relocation in 2016. Most of the trees in the rest of the adjacent mainland were the Chinese Cypresses, used exclusively as nest trees by the herons in 2017-2021.
Distances from the latest used/current nest site (the mainland grove) to the nearest river course (Panlong River, mean width = 30 m) and to the nearest urban lake (Green Lake, 15 ha in water area) were about 300 m and 720 m, respectively.
Survey Methods
From the time of colony formation (~2011), up to 2016, each year around the peak of the breeding season (mid-May), observations were carried out by YQL, who estimated the total number of nests and noted the heron species occupying the nests. Since the colony relocation to the mainland cypress grove (in 2017) (Fig. 1) — until 2021, nest counts were performed on individual nest trees, with the total number of nests calculated. Here we assume that the heron colonies observed before and after the relocation events were the same, based on the geographical closeness of the nest sites used. Activities by humans at the mainland nest site, which conflicted with the herons’ nesting activities (for example, commercial activities, cleaning by zoo staff of fallen items, etc.) were also documented.
Information on new heron species joining the colony, i.e., their annual statuses and numbers, was obtained via interviewing photographer-birders (see Acknowledgments), who had spent extensive time observing herons at the study site.
Results
Colony growth, relocation and new species joining the colony (with or without nesting) are summarized in Table 1. Details on the colony and major human activities at the study site are presented in Table 2. Colony formation, marked by nest-building by Night Herons, took place in two trees at the mainland area in 2011. Since then, Night Heron was found nesting in each year, followed by Little Egret which began to nest in 2012. Several other nests were also recorded with unidentified occupants. Annual nest abundance of the colony — contributed mainly by Night Heron and Little Egret — more than doubled from 2017 (163 nests) to 2021 (418 nests). Colony relocation took place several times, but mainly from the trees at the mainland area to two pond islands, then back to the mainland to the small grove (Fig. 1). Trees on the pond islands were heavily degraded in 2016 (Fig. 2), and the herons moved to the mainland grove to nest, which has more trees with many human activities (Fig. 3).
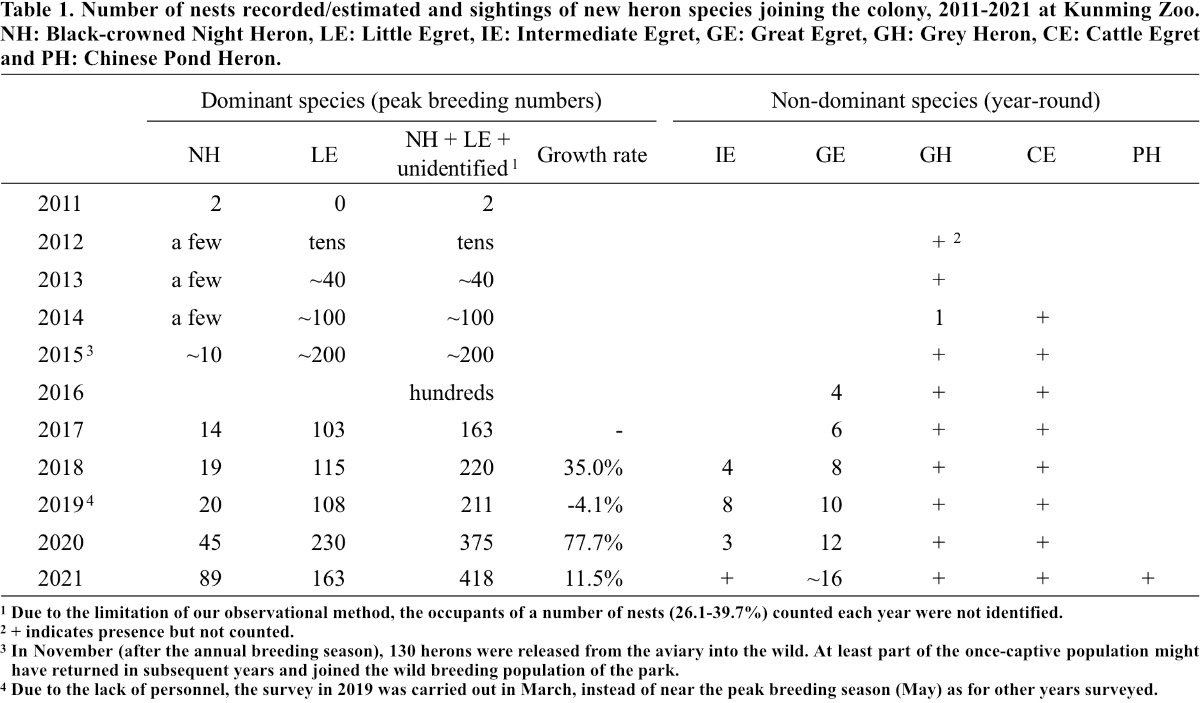
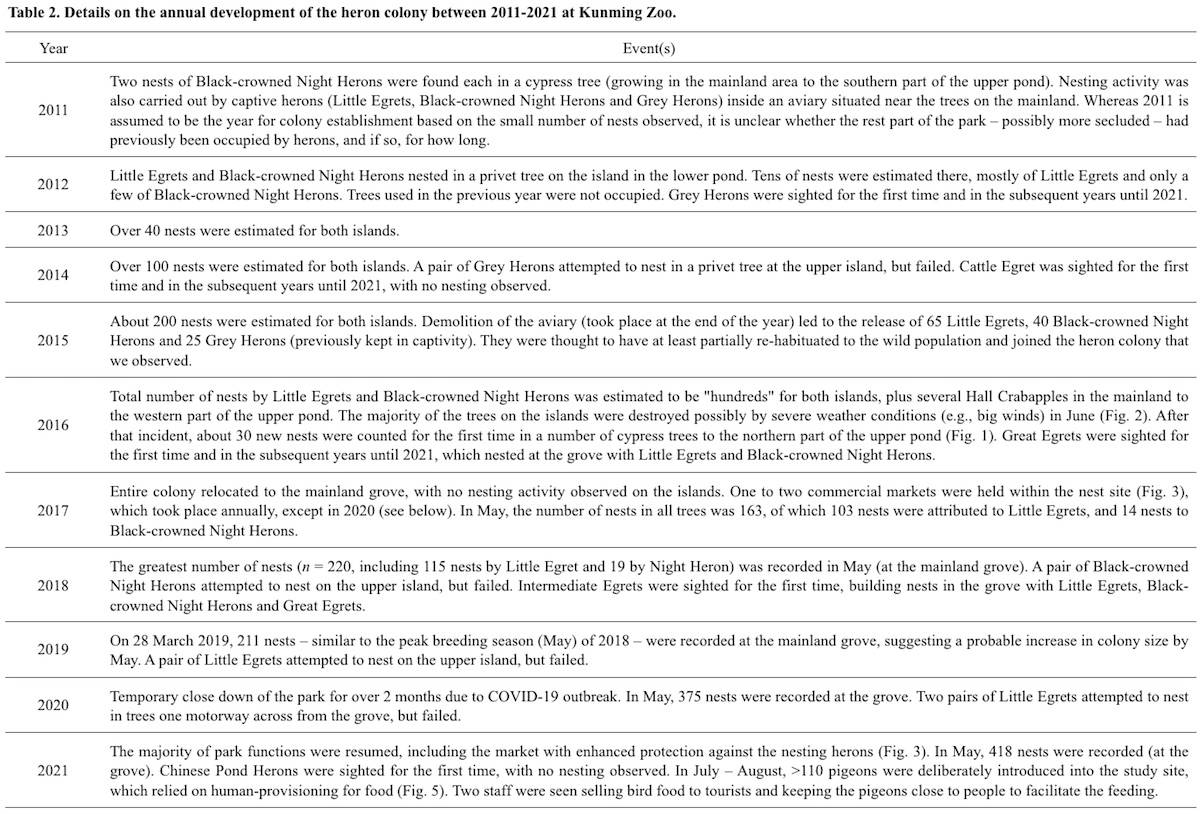


Great Egret and Intermediate Egret were not found nesting until 2016 and 2018, respectively. Nesting by Grey Heron was only observed in 2014. The majority of the non-dominant heron species in the park did not nest (Cattle Egret and Chinese Pond Heron) or contributed very little (Grey Heron, Intermediate Egret and Great Egret) to the observed annual nest abundance. However, nest numbers of Great Egrets, though low, increased annually from four nests in 2016 to 16 in 2021 (Table 1).
Between 1997 and 2015, an aviary (measuring about 13.2 m high and 0.08 ha in area) designed for water birds had been situated near the nest trees that Night Herons used in 2011 (Fig. 1). Three species of herons including Little Egret, Night Heron, Grey Heron and several other species of water birds were kept in the aviary (Fig. 4). The aviary was then demolished on 16 November 2015 and the herons that had been kept were released on the spot into the wild.

Conflicts with human and the COVID-19
Each year during the breeding season, park staff were seen cleaning bird remains, egg shells, prey items (mainly fish, few amphibians), and other particles that had accumulated beneath the nest trees of the grove. Animal and other fallen items were collected manually or were gathered using brooms, and feces were washed off the surface of the concrete floor using a high-pressure water gun and/or common rubber pipes.
At the mainland nest site from 2017-2021, nesting by herons conflicted with one to two annual commercial markets held at the grove, i.e., the herons nested directly above many of the market facilities. During the market years (2017-2019 and 2021) with food-selling activities taking place in a number of booths, the amount of fallen feces produced by herons led to negative hygienic concerns. Protective nets resembling the black sun-shading net with small-mesh size(s) (e.g., 1.7-7.0 mm, Castellano et al. 2008) were set up, which helped to reduce the amount of feces reaching the ground and impacting the pedestrians. During the 2021 market from 07 March to 05 April, more extensive and high-positioned nets (up to 3.1 m high, Fig. 3) of up to about 25% of the tree height (on average 12.6 m) and ~40% of the lowest-positioned nests (by Little Egret, R. Dai unpublished data) were used. The construction process, including the positioning of the nets near the nest trees, could have introduced noticeable impact on the birds, especially those nesting in the lower strata of the canopy.
In early 2020, following the global outbreak of COVID-19 (from late 2019), the park was temporarily closed down from 26 January to 30 March (for 64 days). Tourism was excluded and subsequently, after the park re-opened, restrictions were imposed on the number and size of public aggregations (National People’s Congress, the People’s Republic of China 2007) including at the markets.
Apart from the markets, in July 2021 more than 110 Domestic Pigeons (Columba livia) were introduced to the market grounds below some of the heron nests for entertainment purpose. A small cargo-hut was also established for selling bird food to tourists attracted by the pigeons (Fig. 5).

Discussion
At Kunming Zoo, growth of the heron colony — both in terms of size and of species composition — could reasonably be attributed to several potentially positive factors: increased availability of nest-supportive trees following colony relocation, captive conspecifics in the aviary may have attracted wild individuals from the surrounding area to inhabit the park (Lyles 2000), possible nesting of individual herons released from the demolished aviary as well as stable food supply from provisioning, and reduced predation (Roshnath et al. 2019). Nesting in trees (no nesting was seen in other vegetation categories, such as shrubs) previously not utilized by the herons may indicate colony growth and selection for new site(s), should the current site become less optional for future colony survival and development (Reynolds et al. 2019).
With the expansion of the colony, we have also noticed that trees heavily used by the herons over the years, and covered with feces, seem to have difficulty regenerating, indicating possible habitat degradation in the mainland grove. In addition to some of the major water features within the park area — as potentially important foraging/feeding grounds — the herons may also rely largely on the fish provisioning intended for the captive birds inhabiting the ponds (R. Dai, unpubl. data).
On the other hand, lack of alternative habitat choices in other parts of the city, or changes taking place in colonies outside the park region, may have forced those colonies to move into the park for nesting Such a move could have contributed to the observed increase in the breeding and non-breeding population. However, we do not have direct measurement of these factors.
Whereas the COVID-19, with its huge impact on the human social activities, seemed to have provided an unexpected opportunity for the herons to expand their colonies, increased nesting activities may then have furthered human-wildlife conflicts following the resumption of the park functions. The continued expansion of the colony and increasing heron-human conflicts were thought to lead to a point where colony survival could be jeopardized. This expansion of heron colonies might also cause damage to the limited available nest trees leading to degradation of habitat quality and the possibility of eventually converting the nest site into an ecological trap (Isaksson 2018). In our study area, recently several trees — most of which were situated at the colony center — were heavily covered in the feces by herons, which could lead to negative impact on growth and branch-regeneration even after the breeding season is over.
It would thus be meaningful to monitor the population and dynamics of geographically adjacent colonies within a city landscape (e.g., Kelly et al. 1993), and to document the developmental trajectories of the urban colonies. Those colonies may be very vulnerable to stochastic events, such as the COVID-19, and investigating such situations will further our understandings of human-wildlife interactions and provide scientific basis for preserving species-rich colonies.
Currently there are a number of other Chinese cities (e.g., Beijing Zoo, N. Zhang, pers. comm.; Guangzhou Zoo, J. Mo, pers. comm. and Chongqing Zoo, M. Teng, pers. comm.) reported to have urban parks which serve as breeding and/or roosting habitats for the herons. Those parks, especially those located in highly-populated areas, are in conflicts with the more modernized land-use types and are threatened by a number of relocation initiatives (Liu 2004). However, it would be important to identify some of the old park environments as potentially key-habitat for heron conservation, and to advocate conservation efforts aiming at protecting them from being eliminated.
Acknowledgements
We are thankful to: Peiren Ding, Jianfa Li, Kaiyuan Shao, Jiaxin Song, Kangyin Zeng and Xuepei Zhang who contributed knowledge on new heron species joining the colony; to Ningxin Zhang, Jiaqi Mo and Mingsheng Teng for offering information on herons in other Chinese zoos; to Chao Zhao who helped with species identification; and to Nan Dai, Rui An, Zhentao He who assisted with nest count; and to anonymous reviewers for valuable suggestions made on the English language and contents of the manuscript.
Literature Cited
Angold, P. G., J. P. Sadler, M. O. Hill, A. Pullin, S. Rushton, K. Austin, E. Small, B. Wood, R. Wadsworth, R. Sanderson and K. Thompson. 2006. Biodiversity in urban habitat patches. Science of the Total Environment 360: 196-204.
Carrasco, L., Y. Toquenaga and M. Mashiko. 2017. Balance between site fidelity and habitat preferences in colony site selection by herons and egrets. Journal of Avian Biology 48: 965-975.
Castellano, S., G. S. Mugnozza, G. Russo, D. Briassoulis, A. Mistrotis, S. Hemming and D. Waaijenberg. 2008. Design and use criteria of netting systems for agricultural production in Italy. Journal of Agricultural Engineering 39: 31-42.
Dusi, J. L. and R. D. Dusi. 1987. A thirty-four-year summary of the status of heron colony sites in the coastal plain of Alabama, USA. Colonial Waterbirds 10: 27-37.
Fedriani, J. M., L. V. García, M. E. Sánchez, J. Calderón and C. Ramo. 2017. Long-term impact of protected colonial birds on a jeopardized cork oak population: conservation bias leads to restoration failure. Journal of Applied Ecology 54: 450-458.
Hall, D. I. 1985. Dispersal of a heron-egret rookery. Proceedings of the Second Eastern Wildlife Damage Control Conference. 3-6. [online] Accessed 14 July 2021.
Isaksson, C. 2018. Impact of urbanization on birds. Pp. 235-257 in Bird Species: How They Arise, Modify and Vanish. Fascinating Life Sciences (D. Tietze, ed.). Springer, Cham, Switzerland.
Kelly, J. P., H. M. Pratt, M. Greene and P. L. Greene. 1993. The distribution, reproductive success, and habitat characteristics of heron and egret breeding colonies in the San Francisco Bay area. Colonial Waterbirds 16: 18-27.
Kushlan, J. A. 1993. Colonial waterbirds as bioindicators of environmental change. Colonial Waterbirds 16: 223-251.
Liu, S. M. 2004. On the choice of the way out of China’s urban zoos. China Tourism News 19: 19-24.
Lyles, A. M. 2000. Captive populations, Pp. 293-310. in Heron Conservation (J. Kushlan and J. Hancock, eds.). Academic Press, San Diego, U.S.A.
Mashiko, M. and Y. Toquenaga. 2018. Site fidelity in lineages of mixed-species heron colonies. Waterbirds 41: 355-364.
National People’s Congress, the People’s Republic of China. 2007. Emergency Response Law of the People’s Republic of China. [online] Accessed 29 October 2021.
Nisbet, I. C. T. 2000. Disturbance, habituation, and management of waterbird colonies. Waterbirds 23: 312-332.
Rahman, F. and A. Ismail. 2018. Waterbirds: An important bio-indicator of ecosystem. Pertanika Journal of Scholarly Research Reviews 4: 81-90.
Reynolds, S. J., J. D. Ibáñez-Álamo, P. Sumasgutner and M. C. Mainwaring. 2019. Urbanisation and nest building in birds: a review of threats and opportunities. Journal of Ornithology 160: 841-860.
Roshnath, R., K. Athira, and P. A. Sinu. 2019. Does predation pressure drive heronry birds to nest in the urban landscape?. Journal of Asia-Pacific Biodiversity 12: 311-315.
Rutz, C., M. C. Loretto, A. E. Bates, S. C. Davidson, C. M. Duarte, W. Jetz, M. Johnson, A. Kato, R. Kays, T. Mueller, R. B. Primack, Y. Ropert-Coudert, M. A. Tucker, M. Wikelski and F. Cagnacci. 2020. COVID-19 lockdown allows researchers to quantify the effects of human activity on wildlife. Nature Ecology & Evolution 4: 1156-1159.
Shen, J. X., R. X. Li, F. Xu and S. X. Zhang. 2018. The impact of nest trees pruning on the egrets breeding activity. Journal of Dali University 3: 84-89.
Wong, L. C., Y. Gao, H. Chang, F. Zhou, F. Zou, H. Shi, and W. Feng. 2004. A questionnaire survey of ardeid nesting colony distribution in Guangdong, Guangxi and Hainan, South China. Waterbirds 27: 216-223.
Yan, S. J., X. Zhu and Y. W. Yu. 2006. Construction techniques for the city heronries in Central China. Journal of Zhejiang A & F University 23: 697-700.
Zhang, Y. 2001. Discussion on development of tour recreation and resort in Kunming City. Journal of the Korean Institute of Landscape Architecture International Edition 1: 208-212.