James A. Kushlan3692 Grand Avenue, 618, Coconut Grove, FL 33133, U.S.A.;
Abstract
Although many herons species have robust population statuses, the IUCN/Birdlife International Red List recognizes 12 heron species as being threatened. Common risks among some species include: large-size and forest, coastal and island environments. Habitat loss and change are overall the greatest threats to heron species worldwide. HeronConservation, the IUCN Heron Specialist Group, has, for over 40 years, organized and facilitated heron conservation globally. During this period, global, regional, national and species planning have all contributed to heron conservation action. Threatened species requires species-specific conservation action, but much conservation can be accomplished through larger scale efforts that incorporate the needs of herons. An effective strategy has been creating species working groups, which have facilitated the conservation of several species. Contemporary communication systems allow for interaction, information sharing, informal and formal planning to take place in an effective manner.
Herons - birds classified within the family Ardeidae - are as a group nearly cosmopolitan in their distribution in non-polar regions with some species having accommodated notably well to the human-influenced environment. As a generalization, herons are medium- to relatively large-sized, long-legged, long-billed, long-necked birds, distinctive in neck morphology and in having powder down and pectinate toenails, foraging by wading about in shallow water and roosting and nesting communally with one another (Kushlan and Hancock 2005). That said, among the more than five dozen or so species encompassed within the breadth of the group, each such familial generalization is violated in one way or another by one species or another. Morphology, systematics, behavior and biochemistry studies repeatedly confirm herons to be an evolutionarily cohesive group, as has been uniformly recognized for 250 years since Linnaeus referred all herons he knew to his genus Ardea (Kushlan and Hancock 2005). Recent biochemical study has clarified relationships of species within the family, the most intriguing being the identification of species occupying a more basal position (Boat-billed Heron [Cochlearius cochlearius], Agami Heron [Agamia agami] and tiger-herons) contrasted with the remainder of the family (Kushlan and Hancock 2005, Hruska et al. 2023).
The actual number of species of extant herons is, perhaps surprisingly, unsettled. Hancock and Kushlan (1984) recognize 60 species; Kushlan and Hancock (2005) recognize 62; the IUCN/BirdLife International Red List recognizes 65 including the specific distinction of a large Ardea, the Great White Heron (Ardea occidentalis) (International Union for Conservation of Nature [IUCN] 2024). Future biochemical studies are sure to resolve additional presently uncertain relationships; and, it is likely, they will reveal now unrecognized species distinctions, such as among little egrets (Egretta garzetta subspecies) (Hancock and Kushlan 1984), among Butorides, among Ardea herodias and A. cinerea populations, and among several island forms including night herons (Nycticorax) and purple herons (Ardea) (Kushlan and Hancock 2005).
Herons provide great advantages when it comes to their conservation. They tend to be flexible, and sometimes innovative, in their habits and habitat use. Being relatively long-lived, they exhibit life history strategies emphasizing adult survival such that successful nesting need not occur on an annual basis. This life-history strategy also means that life experience counts. Knowing the local landscape and its seasonal changes, willingness to try out novel options and observational learning are mechanisms allowing accommodation to seasonal and environmental change, individual survival and population resilience. As a result, 81% of the 65 extant heron species recognized by the Red List currently are not considered to be under threat of extinction (IUCN 2024). As an example, Cattle Egrets (Bubulcus ibis), a poster-child example of heron adaptability, has massively increased their ranges within the last century by taking advantage of the human-shaped landscape. Global warming is expediting the long-term trend of heron species’ inching their ranges poleward.
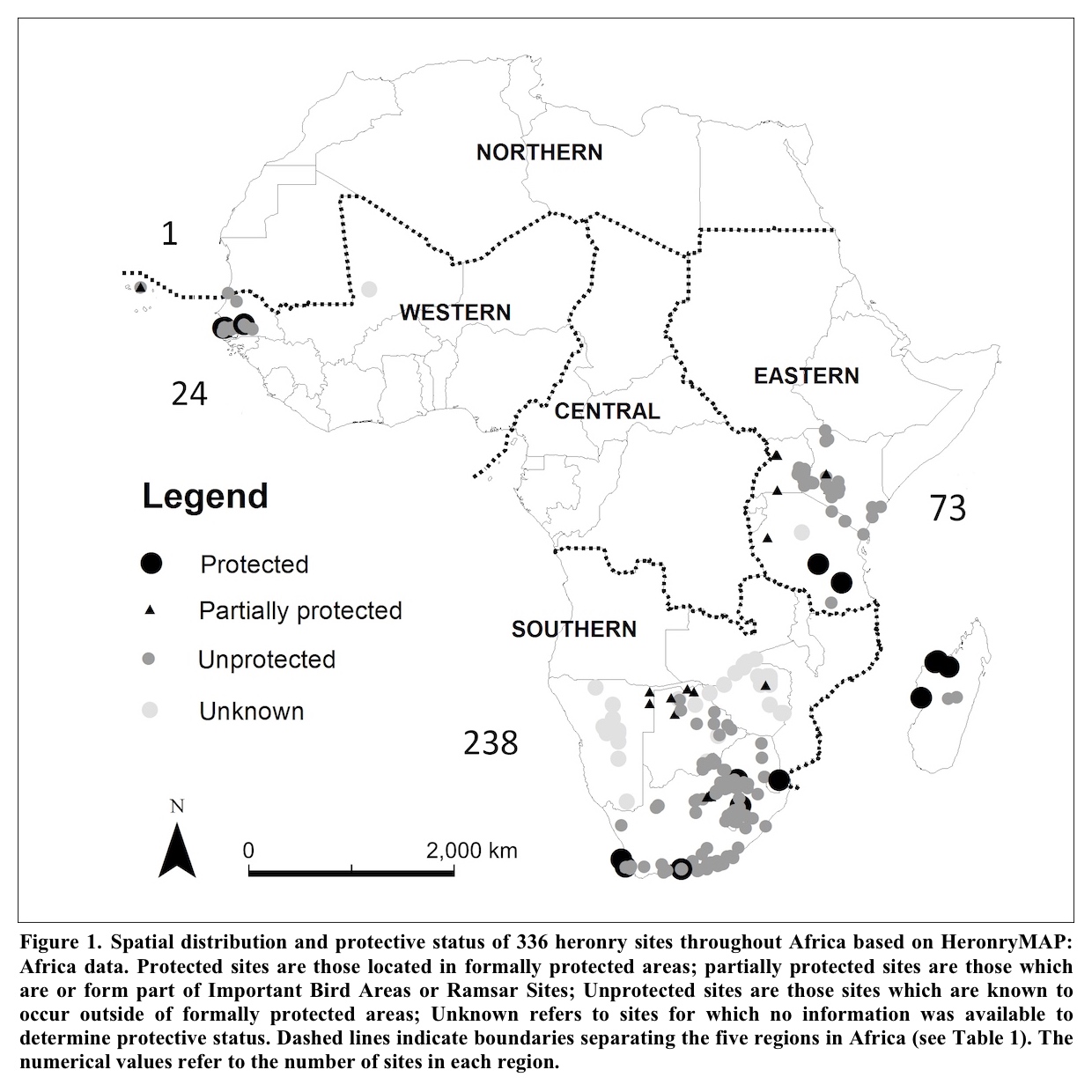
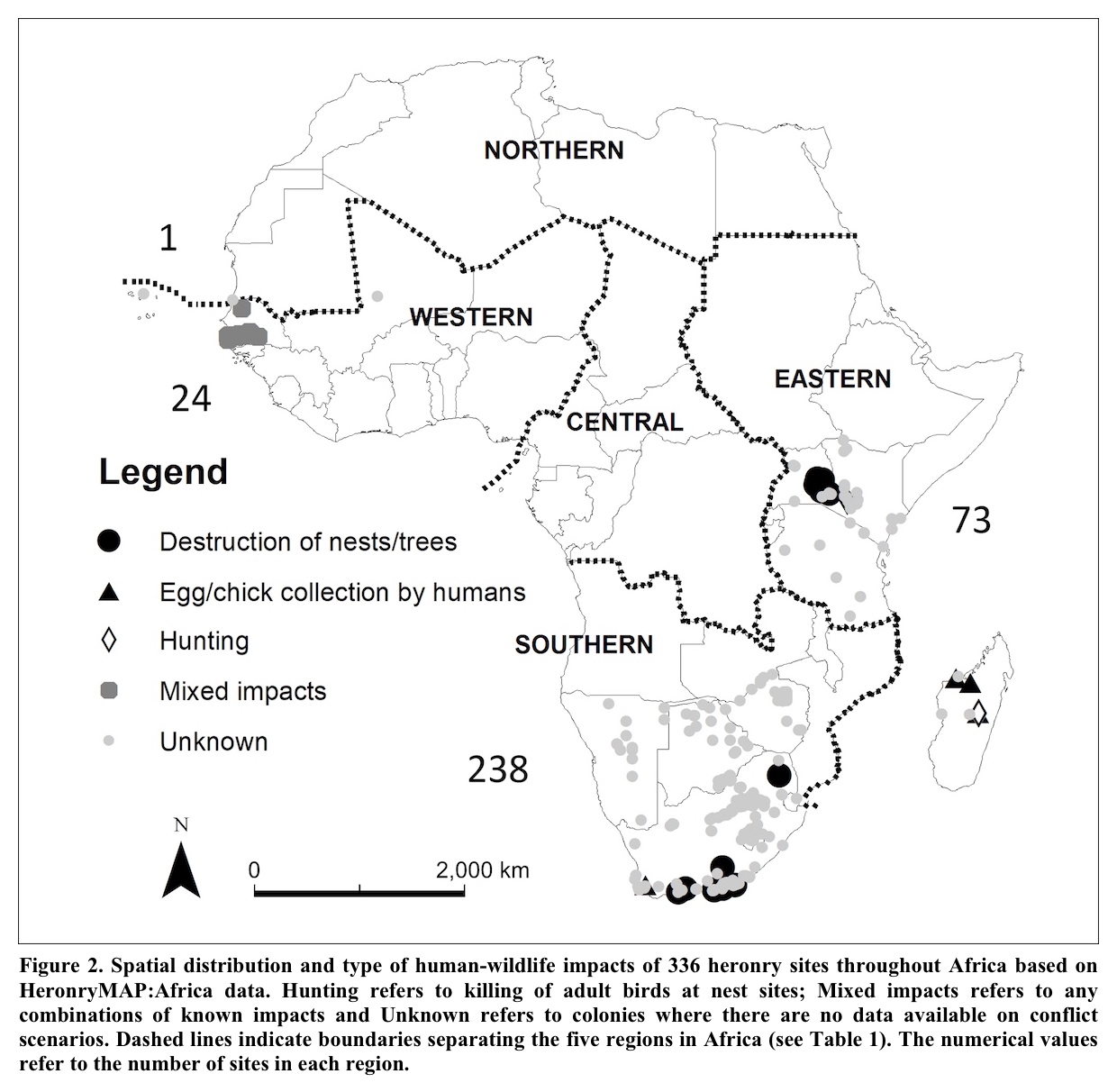
Often locally-abundant and culturally-appreciated, even common herons are not without their conservation challenges. Continued and enhanced protection of nesting areas, feeding habitat, migratory stopover and wintering grounds, as well as cessation of artificial mortality from hunting, egging, nuisance control and accidents all remain required concerns for local heron conservation action. Further, specific populations of otherwise widespread and overall secure species face threats to their stability or persistence locally or regionally. However, the resiliency of the preponderance of heron species and populations allows much of their conservation to be undertaken through broadly encompassing conservation practices rather than requiring species-specific interventions. Depending on locality and opportunities, such programs might include assuring the needs of local herons are incorporated into regional, landscape, and national policies for wetland and woodland management, protective countryside regulations, parks and protected areas, and site-specific conservation initiatives. Such initiatives could include identification and management of Wetlands of International Importance, shorebird reserve networks, flyway initiatives and Important Bird and Biodiversity Areas. HeronConservation has identified 1,723 Important Bird Areas across the globe that are of importance to herons (HeronConservation 2024). Appropriate conservation and management of these areas would accomplish much toward keeping common herons common.
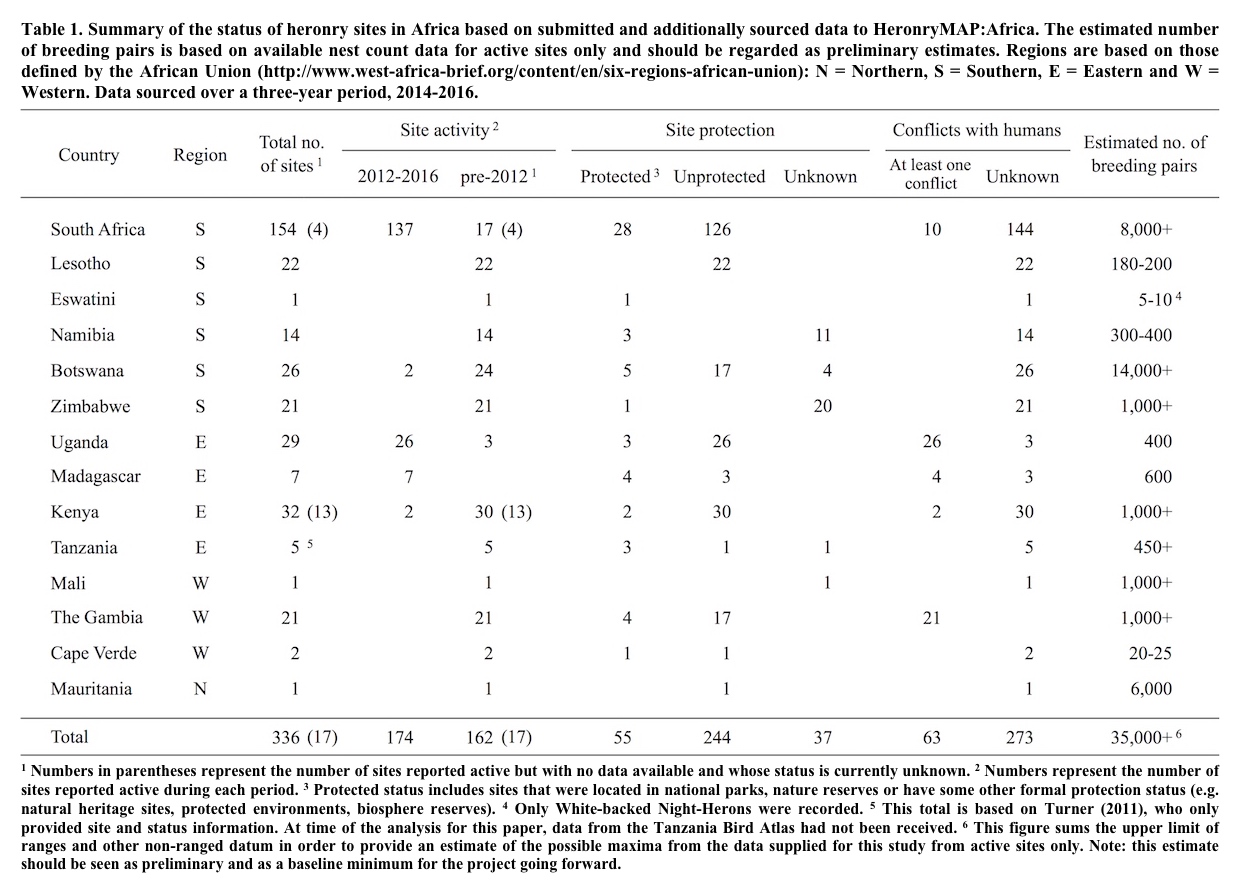
Despite the overall well-being of and resiliency displayed by most of the heron species of the world, a few species are not so well positioned. The IUCN/BirdLife Red List recognizes 12 heron species as Near Threatened or of higher conservation concern with one being Critically Endangered (Table 1) (IUCN 2024). Unfortunately, assuring these species do not become extinct cannot rely on the inherent adaptability of herons nor only multi-species or landscape-scale initiatives; the continued existence of these species requires direct conservation action through active, focused engagement, most effectively led by species champions and stakeholders who successfully engage pertinent conservation, scientific, institutional, political and local communities. Fortunately, research and conservation action in recent years has found approaches that provide hope for the future of even these most threatened of herons. As a result, it is quite possible, despite the drastic situation of a few species, that no extant heron species need go extinct. The objective of this paper is to share some of these approaches, successes and ongoing concerns.
Table 1. Heron species of greatest concern.
Organizing Heron Conservation
Conservation of herons, as is the case for nearly all conservation action, fundamentally depends upon on-the-ground, locally-based engagement protecting breeding sites, managing feeding habitats and inhibiting artificial mortality. These actions are often effectively led by individuals and organizations that become advocates, champions and organizers for the species’ cause. Nonetheless, organization, planning and facilitation above the local scale has proven valuable, perhaps essential. The Heron Specialist Group, organized in 1982, has taken the lead in sponsoring and participating in initiatives at regional and global scales to facilitate heron conservation on the ground (HeronConservation n.d.a).
The most encompassing planning scale is global and is encompassed in the heron global action plan, Conserving Herons (Kushlan 2007). The text of this published conservation plan is now readily available on line (HeronConservation n.d.b). In order to provide the background for this plan, both the biology and the conservation needs of herons needed to be synthesized on a global scale. This was provided by the monographic publications, Heron Conservation (Kushlan and Hafner 2000) and The Herons (Kushlan and Hancock 2005). The specialist group’s publication, Journal of Heron Biology and Conservation, has provided a means for updating these understandings of the biology and conservation of herons.
Tracking the conservation status of heron species and populations is needed to identify changing needs and priorities. Conserving Herons provided a population level analysis of conservation status of the herons of the world. Since its publication, the HeronConservation community regularly participates in updating the conservation status of herons at the species level through the IUCN/BirdLife Red List process (IUCN 2024). The Group also has assembled standard techniques for studying, inventorying and monitoring herons, making this information available on its website.
The values such global-scale planning and evaluation include: providing a common base of understanding, global perspectives, shared strategies and tactics, common nomenclature and identifying priority actions. Also, by identifying priorities from an international perspective, a global plan can be used to justify and validate specific efforts that might be proposed to meet these priorities. New information, new opportunities and new priorities are continuously being uncovered. Updating global action plans and other global information need not require a committee or hard-copy publication but rather can be accomplished continuously through open-sourced volunteered information from specialist group members.
The next stepdown in planning scale is at the multinational scale. For most species, this would occur within the context of multispecies planning such as regional waterbird plans and flyway-scale initiatives. Heron conservationists must be certain to be involved in such initiatives so that herons are “at the table” when needs, priorities and funding are being negotiated. In the Americas, such regional planning for herons was encompassed within the North American Waterbird Conservation Initiative that resulted in a multinational waterbird conservation plan, Waterbird Conservation for the Americas, which brought herons among other waterbirds into the ongoing evolution of bird conservation in North America (Kushlan et al. 2002, Kushlan 2012). It is crucial for the needs for heron conservation to be incorporated into opportunities provided by international flyway initiatives, such as the African Eurasian Migratory Waterbird Agreement, the East Asian-Australasian Flyway Partnership and Regional Flyway Initiative, and the Americas Flyway Initiative.
A crucial scale for heron conservation planning and implementation is national and subnational because this is the scale at which governmental conservation priorities are set, organized administratively and funded – if they are to be funded. It is also the scale at which laws and policies determine much of what heron conservation can hope to accomplish. Engagement of heron conservation is essential if the needs of herons are to be appropriately considered within the conservation framework of a nation, state or province. National planning follows on the global planning and evaluation models, i.e., first documenting the distribution, population size and conservation needs of species of herons found within a country to the extent practical. This should be followed by evaluating the appropriate relationships of heron conservation needs within national conservation regulations and processes, and identifying priority actions. Such national syntheses of biology, status and needs are essential first steps, reviews for Ecuador and Paraguay are recent examples (Cisneros-Herdia and Penaherrera-Romero 2022, Yanosky et al. 2023). Publication of peer-reviewed national analyses of heron status and conservation needs remains a high priority so as to establish accepted population bases for conservation action within a nation.
Some species require species-specific planning at whatever scale is appropriate. Detailed species-level planning for each threatened heron species, preferably set within the context of global and regional planning and implementation, is essential as conservation action generally needs to extend beyond the benefits provided by multispecies and regional initiatives. Species planning and implementation of conservation measures can be markedly effective because they are specific to the most pressing needs of a species; they pinpoint the most important do-able actions, identify important sites for protection and management, and enlist engagement and commitment of government, organizations and species champions. In some cases, a species can become recognized as an indicator of environmental health, valued as a natural patrimony of a country or a symbol of conservation action such as White-bellied Heron (Ardea insignis) in Bhutan and Great Egret in the United States. Because such action is species- and geographically-specific, it can take into account national and local regulations and opportunities. Because such action addresses the most pressing of issues, it can be efficient and cost effective.
Several decades of experience has shown that species planning takes place most effectively through a working group process. Species-specific heron working groups have arisen in various ways. Several such working groups are recognized and facilitated by HeronConservation. Other approaches also have been used such as being organized by an individual, organizations or funding entity. Whatever works for a species is welcomed. In all cases, partnerships and partnership groups are generally necessary as part of recovery implementation.
The working group process generally begins with discussions among those individuals having a concern for the wellbeing of a species, followed by one or more meetings to gauge interest and potential for concerted conservation action. If interest is sufficient, a decision is taken to form into a group to facilitate ongoing engagements. The first order of business, if necessary, is to work together to develop a better understanding of the distribution and population size of the species. At the same time, to the extent possible for each species, coordinated and collaborative research carried out by group members attempts to fill gaps in knowledge essential to conservation action. Continued communication and interaction among participants are essential, made more feasible through electronic communications. Communication linkages build the network of participants eventually, hopefully, to encompass the species range.
The collaborative development of a first-version species action plan serves to clarify information gaps, assign responsibilities, identify conservation needs throughout the range, create monitoring programs, identify potential obstacles to recovery, provide measurable implementation goals and timelines and serves as a process to enhance group engagement and cohesiveness. As the group matures, understanding increases and conservation action gets underway; the plan should be updated regularly. Subplans might be created on geographical, political or subpopulation bases. A crucial part of the planning process is to identify financial resources available, whether through targeted grants, organizational commitments or government sources. A business plan as part of or separate from the species action plan may be used to identify how organizing and funding the recovery is to be done. Grants are usually needed but need not accrue to the working group itself but rather to individuals and institutions working within context of the group’s plan. There is little reason for a working group to incorporate or create an independent charity as these roles can be played more efficiently by partner organizations. The important thing is that funding get to the ground level by whatever mechanisms are available, from whomever can deliver it, to whomever can achieve its goals, and that the funded activity be successfully carried out, evaluated and reported. Throughout the process, one of the most important roles of the working group is to maintain positive communication among its members and partner organizations. Maintaining the group can sometimes be harder than forming it, as it takes persistent leadership, engagement and successful fundraising.
Threats in Common
Twelve heron species are of most immediate concern at present (Table 1). They share characteristics associated with their threatened status. For most, of course, threats come in more than one variety and there is generally more than one threat to a threatened species persistence. Nonetheless some communalities emerge.
Size counts. Several species of Ardea are exceptionally large birds, including the Critically Endangered White-bellied Heron, second in size only to the Goliath Heron (Ardea goliath), which can stand 150 cm tall; and an even larger heron was extant until about 5,000 years ago (Kushlan and Hancock 2005). Although this great heron of the Middle-east went extinct, likely by being eaten by humans, the African Goliath Heron is relatively secure at the moment. But life as a large heron is not easy. In feeding, there needs to be a tradeoff between the energy used to catch prey and energetic return; so large herons tend to eat large and relatively infrequently-encountered fish. It is not unusual for a large heron to capture one or two fish a day. Standing around waiting for a large fish to come by is always tenuous. Also being large means that the nesting period is long, as the chicks need time to grow to fledging size. The longer the nesting period, the greater the risk of failure in any one year.
Two herons of conservation concern are large, the White-bellied Heron and the Great White Heron. They live in difficult foraging situations, the White-bellied Heron using forested wetlands especially rapidly-moving mountain streams and the Great White Heron using tropical marine shallows having clear water and nowhere to hide from their prey. Large herons are not so mobile as smaller ones, so they tend to have a limited repertoire, standing and waiting for a suitable fish to come by. These large-bodied heron species also represent large, rather sedentary and approachable food source for people. Grey Herons (Ardea cinerea) were featured dishes in Medieval Europe (Kushlan and Hancock 2005).
Foraging at night also has its challenges, requiring both evolutionary adaptation and successful application of learned skills. Two species of concern are night herons, the White-eared Night Heron (Oroanassa magnifica) and the Japanese Night Heron (Gorsachius goisagi). Night herons are among the most widespread herons. As might be expected, night foraging is not all that easy. Other night heron species (Gorsachius, Oroanassa, Calherodius) are range-restricted and inhabit breeding and wintering areas subject to deforestation as well as other threats - the persistence of the White-eared Night Heron in southern China, for example, was confirmed by a bird being offered for sale at local market (Kushlan and Hancock 2005). Island and other isolated forms of night herons are more at risk than the official listings suggest; and so, additional biochemical studies are needed to determine their taxonomic and conservation status.
As suggested in the examples above, forest-dwelling heron species face challenges. Four species of concern are forest-dwelling herons, White-bellied Heron, White-eared Night Heron, Agami Heron and Forest Bittern (Zonerodius heliosylus). These species depend not only on forests but on availability of specific aquatic habitats available within the forests. Documenting the conservation status of forest species is often problematic as inventory and censusing are so difficult over broad stretches of forest habitat. This uncertainty can lead to disagreement in opinions as to a forest species conservation classification depending on whether one gives more or less credence to the documentable population size or to the amount of potential habitat available to the species. Population status determination based on known nests or individuals are likely underestimations whereas those based on extent of possible habitat are likely overestimations. So, in part, the official status of forest species of concern may be the result of limited data. Or, alternatively, the massive pattern of forest alteration – previous, underway or impending – around the world could not help but be potentially devastating to populations. It is without question that these birds are threatened by such rampant forest conversion on both breeding and non-breeding grounds.
In addition to forests, two other environments emerge associated with some heron species of concern, those using coastal environments and those restricted to islands. Tropical marine environments are particularly tricky for visually-feeding herons, as successful foraging in shallow, warm, clear water in the bright sunlight is not all that easy; the fish can see them coming. Chinese Egrets (Egretta eulophotes) nest and winter along the coast and Great White Herons and Reddish Egrets (Egretta rufescens) must deal with this exposed, low-productivity environment year-round.
Islands too pose challenges. The Black-crowned Night Heron (Nycticorax nycticorax) and its close relative the Rufous Night Heron (Nycticorax caledonicus), between them, are found on all the inhabited continents and many islands. However, this dispersal ability has also led to isolation on islands. Four of the five heron species that have recently gone extinct were island populations of night herons: the Mauritius Night Heron (Nycticorax mauritianus), Reunion Night Heron (Nycticorax duboisi), Rodrigues Night Heron (Nycticorax megacephalus) and Bermuda Night Heron (Nyctanassa carcinocatactes) (IUCN 2024). The fifth, New Zealand Little Bittern (Ixobrychus novaezelandiae) was also an island species. Japanese Night Heron, Malagasy Pond Heron (Ardeola idae) and Madagascar Heron (Ardea humbloti) are similarly island forms at risk (IUCN 2024). Heron species and subspecies (including an endemic race of Purple Heron (Ardea purpura madagascariensis) in Madagascar face degraded habitat as well as hunting. The Malagasy Pond Heron winters in Africa, so habitat availability in this area is also of concern.
Marsh-dwelling bitterns, particularly species of the great bittern group, Botaurus, tend to be at risk. Each of the four species of great bitterns are in fact of long-term conservation concern facing challenges throughout their ranges. These species depend on thickly-vegetated marshes. Accurate population information is difficult to obtain because censusing such cryptic birds relies on intensive and extensive surveys in wetlands using call detection. Fortunately, large bitterns call mightily. The heron species of concern include a species of bittern, the Australasian Bittern (Botaurus poiciloptilus). Given that these birds are dispersed and territorial, significant areas of marshes having appropriate water conditions and management are needed. The marshes used by the Eurasian Bittern (Botaurus stellaris) are affected by drainage and cane harvest. One population of the Eurasian Bittern (Botaurus stellaris capensis) occurs in a disjunct distribution in southern Africa, and its breeding population status remains poorly determined (Kushlan 2007). Similarly, little is known about the Pinnated Bittern (Botaurus pinnatus) (Kushlan 2007). All of the great bitterns deserve additional study and further evaluation of conservation status of their various populations.
A common theme among heron species of concern is the role of habitat loss, change and degradation. In fact, habitat change is the greatest communality among the species known to be at risk. Some species such as White-bellied Herons, Great White Herons and Slaty Egret (Egretta vinaceigula) have limited habitat and geographic breadth and so are dependent on having appropriate conditions in relatively limited space. As noted above, forest-dwelling species are faced with forest clearing and conversion; great bitterns rely on very specific reed marshes that, depending on location, face threats from drainage, conversion, exploitation and climatic change. Madagascar may be an epicenter for wetland habitat-change threats to herons of several species and subspecies. Thirty species of herons have been evaluated as decreasing; nearly all have suffered some aspect of habitat limitations (Kushlan 2007). Along with nesting site protection, habitat management that favors herons is an overarching need for conserving herons globally and locally.
Threatened Herons
White-bellied Heron - The heron species of greatest concern, globally, is the White-bellied Heron of Asia, the only heron currently-listed as Critically Endangered. Once called the Imperial Heron (Ardea imperialis), its known nesting pairs can be counted on one hand, and these counts vary frighteningly from year to year. In Bhutan, 1 to 5 nests have been found annually with the population there estimated to be between 14 and 30 individuals (Royal Society for Protection of Nature [RSPN] 2022); the total world population is estimated to be fewer than 60 birds. As a long-lived heron with such a small population size and wide dispersion, any adult mortality or nesting failure, artificial or otherwise, can have a significant global population impact. Its ecological situation is precarious as it only occurs in the foothills and lower elevations of the Himalayas to about 1,500 meters. It is a wary, easily disturbed bird, nesting singly in tall trees and foraging in undisturbed ponds, lakes and fast-flowing rivers, all of which are highly vulnerable to man-made and natural disturbances including water power development, gravel mining, pollution and hunting (e.g., Khandu 2022). In Bhutan, a national priority is the development of hydropower on its mountain rivers, which would affect habitat both upstream and downstream of dams. Encouragingly, recent results show it can at times habituate to human presence (Patgiri 2022).
As of the mid-1980s, only two nests had ever been reported (Hancock and Kushlan 1984). The species nesting was essentially rediscovered in 2003, with a report of a nest in Bhutan. Over the next two decades, the desperate situation of the species led to the development of an international conservation effort (Acharja 2022, Bida 2022, Goodman 2022). A key element of this initiative was the emergence of species champions, especially the Royal Society for the Protection of Nature in Bhutan, which began study and conservation measures soon after the 2003 discovery (RSPN 2019). Other crucial champions and partners joining over the following decades included Bhutan civil authorities, Bhutan Trust Fund for Environmental Conservation, the Bhutan hydroelectric sector, non-governmental organizations, and foundations, notably the leadership of Gemma Goodman of the British-based Synchronicity and George Archibald of the International Crane Foundation, and the support of the MAVA foundation (RSPN 2019).
Meetings of interested parties from across the species’ range took place in 2014 and 2015 that led to creating a working group and developing a conservation strategy (Price and Goodman 2015, RSPN 2019, 2022, Goodman 2022, Royal Society for Protection of Nature and Department of Forests and Park Services 2022). As a result, understanding and acting on the needs of the species were increasingly engaged especially in the species’ apparent stronghold in Bhutan. These actions included enhanced survey and monitoring, experimenting with research and monitoring techniques, expanding work in India and China, initial attempts at captive rearing and movement studies. The working group concluded that, given the species’ extremely precarious status, undertaking the drastic measure of a captive breeding program was essential, requiring partnerships among governments, scientists in different countries, the zoo community and private industry. With this partner-rich support, the breeding center was built in 2020 and the first chicks introduced there in 2021 (Royal Society for Protection of Nature and Department of Forests and Park Services 2022). Meanwhile inventory, research and monitoring continue in nearly all range countries and the specialist group continues to provide connectivity. Perhaps the most pressing need at this moment is to secure additional and continuing funding for the captive rearing center, including establishing an endowment. Continuing inventory, monitoring, habitat use and migration research across the species range remain priorities. Securing continuing institutional support for the activities of the working group is essential.
White-eared Night Heron - This night heron, restricted in range to southern China, northern Vietnam and northern India, was for years little known and was evaluated as Critically Endangered in 1994 (Collar et al. 1994, Pilgrim et al. 2009, Shafli 2018). The Heron Specialist Group early identified the species as high priority, raised concerns, and participated in surveys and status reviews (Fellowes et al. 2001). Fenqi He and colleagues later documented that the species habitat was far more widely distributed in China than previously thought including in protected areas and the range may not be as fragmented as then believed (He et al. 2007, 2011). The species conservation status was re-evaluated and, with an estimated world population of under 1,000 in 2000, the species was determined to be endangered (BirdLife International 2017a). The species is legally protected in China, although not in Vietnam. Local food hunting does occur. Its habitat is montane and sub-montane subtropical and tropical forest and associated rivers and marshes but also it is recorded in plantations and secondary-growth pine forest. Despite apparently showing some level of accommodation in habitat use, its principal threats remain related to habitat change and forest management including timber harvest and conversion to plantation planting and other forms of agriculture.
The species does not yet enjoy a global conservation action plan, although a local plan has been prepared in Vietnam (Dine 2012). Conservation action for this species requires an increase in the breadth of surveys, monitoring and modeling to achieve an accurate understanding of population size, dispersion and trend. Research is also needed to determine connectivity among the known subpopulations so as to understand the extent of the population’s fragmentation. Telemetry studies are similarly needed to determine migration patterns and wintering sites of specific breeding groups. The encouraging recognition of the species, in some areas, as a symbol of forest conservation is welcome, a recognition that should not depend on its achieving any specific conservation assessment classification. Public awareness programs to inhibit hunting are needed wherever this activity may occur; and the species needs to be given official protection in countries where it occurs but is not now legally protected.
Malagasy Pond Heron - This heron breeds in Madagascar and nearby Mayotte, Réunion and Aldabra; it winters in Africa. Classified as Endangered, it has an estimated world population of under 2,500 birds and is declining (Rabarisoa et al. 2020, BirdLife International 2021). Its habitat primarily is freshwater wetlands, lakes, and rivers; although on Aldabra it uses mangrove swamps and shore lagoons (Pruvot et al. 2020, Rabarisoa and Hajanirina in review). In Madagascar, exploitation of its eggs and young and other disturbance to nesting sites remain threats. Foraging habitat is critical and this species has the ability to accommodate to prey variation throughout its breeding period (Pruvot and Rene de Roland 2021). So, in addition to nest site protection, adjacent foraging habitat also requires conservation attention. Overall, its main threats are habitat destruction such as conversion of wetlands for agriculture, deforestation and urbanization. Another threat may be hybridization with the Squacco Heron (Ardeola ralloides) (Betleja 2023).
A conservation action plan for the Malagasy Pond Heron has been prepared. The Madagascar Pond-heron International Working Group has been established under the auspices of the African-Eurasian Migratory Waterbird Agreement (Ndang’ang’a and Sande 2008). The working group is composed of representatives of national authorities in consort with invited experts and conservation organizations. The group’s goals include setting priorities for action and coordinating implementation of the action plan. Important sites have been identified range-wide and the breeding population is monitored. Although this heron’s white breeding plumage is unmistakable, in winter plumage it closely resembles Squacco Herons, which adds difficulty to monitoring its status on its wintering grounds. The existence of an international working group composed of national representatives to oversee implementation of the recovery plan is encouraging. The work of such a fundamentally intergovernmental board might benefit from the conservation-action engagement of non-governmental organizations, alliances, scientists and private individuals either within or adjacent to the formal working group. The conservation issues for herons as a whole in Madagascar require active intervention to conserve and manage wetlands, to assure availability of water supplies for conservation, and to inhibit hunting. Monitoring of the breeding population needs to continue; and movement and telemetry studies to better pinpoint African wintering areas are needed to assure conservation of these habitats.
Madagascar Heron - Madagascar Heron is endemic to Madagascar, although there are records from the Comoro Islands and Mayotte. In Madagascar, it is restricted to the western part of the island. The current population, estimated as fewer than 1,500 individuals and moderately declining, is considered to be Endangered (BirdLife International 2016a, Rabarisoa 2022). Its main habitats are coastal areas and freshwater lakes and rivers. Threats include overexploitation from hunting and egg collection and loss and degradation of its wetland habitats. Colony site monitoring and protection and habitat conservation are crucial actions for the species’ conservation.
As is the case for the Malagasy Pond Heron, a principal conservation concern for the Madagascar Heron is preserving and managing remaining wetland habitat, which also benefits other waterbirds including the endemic race of the Purple Heron. The Madagascar Heron does not yet enjoy the attention of a specific working group or guidance of a conservation action plan. Perhaps as a complement to the government-based Madagascar Pond-heron Working Group, a primarily non-governmental, multispecies Madagascar Ardeidae Working Group might have value.
Great White Heron - This heron’s range is primarily restricted to southern Florida and, to a lesser extent, Cuba (McGuire et al. 2019, BirdLife International 2020a). It inhabits shallow coastal marine lagoons and less frequently freshwater wetlands. Its principal population in Florida, although occurring primarily in protected areas, has been affected by habitat degradation (Powell 1983). This heron has recently, and belatedly, been recognized as a separate species (BirdLife International 2020a, Browning and Kushlan 2022). An important next step in its conservation is better to assess global population size, trend and distribution, especially in Cuba, Mexico, Venezuela and the Caribbean so as to be able to assess genetic structure and determine connectivity among populations.
The most critical action for the conservation of this species is to reverse quickly the deterioration of its principal habitat in Florida Bay, Florida, USA, especially to assure and increase and maintain its food supply. Although the Great White Heron National Wildlife Refuge is dedicated to the species, conservation action has not progressed much beyond monitoring. This is a species in desperate need of a conservation action plan developed by government agencies, conservation groups and scientists for at least the Florida population. A working group would facilitate this accomplishment. Action in Florida should be accompanied by support for a complete inventory of the species in Cuba.
Australasian Bittern - This large bittern has an estimated world population of only 1,500-4,000 birds in New Zealand, New Caledonia and Australia where the population of about 1,300 birds is concentrated in New South Wales (Herring 2022). Although some subpopulations occur in natural wetlands (Fitzsimonds 2022), 500 to 1,000 birds are supported by rice farms (Herring et al. 2019), which has provided an opportunity to connect bittern conservation with rice field management through a heron conservation program called Bitterns in Rice (Bitterns in Rice Project n.d.). The project is a collaboration among the Rice Growers’ Association of Australia, Birdlife Australia and others with significant governmental engagement. A conservation plan has been prepared and conservation meetings are held (Commonwealth of Australia 2022, New South Wales Government 2022). Critical research has provided models for co-management of rice fields in ways conducive to heron conservation to guide incentives for bittern-friendly rice farming (Bitterns in Rice Project 2016). However, despite increased attention, changes in water use management within rice fields provide additional threats to rice-inhabiting bittern populations, as do climate change and increasing droughts requiring the necessity of providing drought refuges (Herring et al. 2021, Herring 2022).
The Bitterns in Rice Project has shown marked success in bringing attention to the bittern and producing methods for co-management of rice fields. However, pressures to change agricultural practices in light of increasing drought can undermine this effort. Government intervention and support and conservation allocation of water supplies will be required. Natural wetlands in which the species nests need to continue to be preserved and managed appropriately wherever the bittern is found.
The several species of great bitterns face challenges everywhere, but conservation successes have also been achieved (White et al. 2006). Increased communication among great bittern biologists and conservation advocates among all the species would prove of great benefit, perhaps aided by the establishment of an international, multispecies great bittern working group.
Japanese Night Heron - This range-restricted species was classified as Endangered in 2000; but, more recent surveys have suggested that its population is larger than previously thought and so was reclassified as Vulnerable (BirdLife International 2020c). However, based on ongoing deforestation across breeding and wintering areas and trapping pressures across wintering grounds, the population is inferred to be undergoing a continued decline. The species nests almost entirely in Japan (with a few records from Taiwan and South Korea), winters in the Philippines and is transitory elsewhere (Kushlan and Hancock 2005, Oh et al. 2010). This species declined starting in the 1970s disappearing from many of its former breeding sites; but recent breeding surveys and records from the Philippines suggest a larger than previously known population (BirdLife International 2020c). The species uses a diversity of forest habitats, from patches to heavily forested for both nesting and wintering (Hamaguchi et al. 2014). It also uses swamps and rice fields. Ongoing deforestation across its range and hunting on its wintering grounds continue as threats and suggest a continued population decline.
The understanding that the species may be more abundant and widespread than previously thought is encouraging. However, modeling should be accompanied by a thorough effort to conclusively document breeding population size and its specific dispersion within Japan and to assure that these sites are protected. Movement and telemetry studies are needed to clarify its wintering grounds and what specific threats to those sites might be mitigated.
Slaty Egret - With its breeding population concentrated principally in Botswana in the Okavango Delta and Chobe River, this heron has a restricted range and habitat availability. The population is considered to be fewer than 3,300 adult birds, declining, and is classified as Vulnerable (BirdLife International 2016c). It nests in seasonally flooded wetlands starting when water levels are beginning to recede from their seasonal peak (Kushlan and Hancock 2005). When not breeding, it disperses widely moving about, apparently in response to rainfall (Hancock et al. 2006). Considerable efforts have been made at discovering colony sites, with known sites in Botswana increasing from 3 to 12 in the past 20 years as have other records reported from outside the nesting season (Tyler 2005, 2024). Although suitable habitat appears to be available, its documented population remains small. A species action plan was published in 2012 (Tyler 2012). However, no working group has been established and so organized follow-up to the plan has been limited. Major threats include wetland conversion to agriculture, disturbance at foraging and nesting sites, impact of non-native floating plants and taking of eggs and fledgling birds.
As the breeding range of the species is relatively limited, the most crucial conservation action is creating permanent protection for more of the Okavango Delta, especially the northern Panhandle. Much of its habitat is accessible only with great difficulty in the wet season. Nonetheless, thorough and systematic monitoring efforts are needed and additional colony sites in Botswana and possibly in other nearby countries should identified and protected. Outreach programs, especially to the tourist sector, are needed to relay information on the impact of disturbance and how to avoid it.
Chinese Egret - This heron nests almost entirely on small, uninhabited islands off the Asian coastline, including mainland China, eastern Russia, and North and South Korea but has been extirpated from Taiwan and Hong Kong (BirdLife International 2016b). It is migratory, wintering in the Philippines and Malaysia (Li 2006). Its population is estimated to be under 10,000 birds and is declining long term, although its population may be stabilizing in China and previously unknown colonies have been discovered. It uses shallow tidal estuaries and bays, mudflats, fishponds and paddy-fields.
The species is legally protected in its breeding range countries, although egg harvest occurs. Its declining status is primarily due to the loss of estuaries and mudflats from reclamation and loss of breeding sites due to industrial, residential and agricultural development. Protecting nesting areas and feeding areas is essential, requiring incorporation of the species needs into regional and local conservation and development planning. Its wintering areas require better definition and monitoring, especially in its apparent areas of concentration in Malaysia and conservation measures enacted in important wintering areas.
Agami Heron - This unique heron nests, sometimes in huge colonies, in Central and northern to central South America but disperses widely in the non-breeding season, sometimes at great distances from the colony site (Reynaud and Kushlan 2004, Stier et al. 2017, Chen et al. 2022). Its nesting and reproductive success depends on the extent of the wet season. Much of its known population nest in a few large colony sites while its foraging areas include wide regions of forested wetlands and streams. Among its other distinctive features are its plumage, its much-elongated morphology and its behavior (Kushlan and Hines 2016). Its forested habitat is under threat and so the species is classified as Near Threatened (BirdLife International 2023). A species conservation action plan has been published and a working group established that facilitates communication, reporting, monitoring, research, and conservation (Stier and Kushlan 2015).
The vulnerability of this species, owing to its nesting in a few large colonies, emphasizes the need for protection of these sites from disturbance, including egging, which has proven to be an effective conservation tactic for the species (Chen et al. 2022). Conservation of its feeding habitats is more difficult, and requires the species needs be considered within large habitat conservation efforts, especially the establishment and management of protected areas and reserves. Continuing engagement of the Agami Heron Working Group is essential as is increased funding and institutional support for its activities.
Reddish Egret - This heron occurs exclusively along tropical and subtropical coastlines from southern North America, through the Caribbean, and Central and northern South America with birds dispersing southward in non-breeding periods. Its restricted, patchily distributed habitat includes shallow lagoons, marine flats and shorelines, and small, protected island breeding sites. Because of its restricted and threatened habitat, it is considered to be Near Threatened (BirdLife International 2020b). An international working group was established in 2006 following an organizational workshop and status assessment (Green 2023). From 2006 to 2011, members of the group conducted coordinated research including genetics, established banding and telemetry programs to assess genetic population structure and movements, conducted inventory studies and established a network of stakeholders. Species conservation action plans were completed in 2012 and 2021, as well as was regional planning (Wilson et al. 2014, Green et al. 2022).
The species is at risk because of its restriction to shallow tropical shores, which are under threat through much of its range. Although many of its known colonies are in protected areas, other important nesting and foraging areas should similarly be protected from development, environmental alteration and disturbance. The working group’s plan (Tarbox et al. 2020) provides a ten-year set of measurable goals for the species and its subpopulations.
Forest Bittern - One of the least known herons, the Forest Bittern, also called the New Guinea Tiger-heron, is restricted to forests of New Guinea (Kushlan and Hancock 2005). Its estimated population is 7,000 or less and is considered Near Threatened (BirdLife International 2017b). But the species is rarely seen or reported so information is really not available on its population size (Pratt and Beehler 2015). However, its threats are clear. Habitat loss, including logging, deforestation and conversion of its forest habitat.
There are few herons for which the information base is so slim. Encouraging increased reporting, formal inventory and census where feasible, and basic biological research all are needed. However, its conservation is clearly linked to the health of New Guinea’s forests and forest streams, which are among the most threatened in the world. Conservation of the species and its forests in Indonesia and Papua New Guinea is a matter of international concern and controversy given limited environmental responsiveness by the government to these threats.
Discussion
Aided by their accommodative abilities and resiliency, most heron species are not at immediate risk of extinction. However, some are, likely including 12 currently listed on the IUCN Birdlife International Red List. Threats to the persistence of the White-bellied Heron, Australasian Bittern, Malagasy Pond Heron and Great White Heron are indisputable and require immediate action. Others, such as White-eared Night Heron, Chinese Egret and Reddish Egret may actually be more secure than presently recognized. Some species, such as Japanese Night Heron, Slaty Egret, Agami Heron and Forest Bittern, suffer from critical information gaps inhibiting appropriate assessment of their conservation status and needs.
The Red List evaluates conservation status at a species level. However, specific populations are also at risk, including the Purple Herons of Madagascar (Ardea purpurea madagascariensis) and Cape Verde (Ardea purpurea bournei), the Grey Heron (Ardea cinerea monicae) of Mauritania, and the Eurasian Bittern (Botaurus stellaris capensis) in South Africa. These and such populations as the Lava Heron (Butorides striata sundevalli) of the Galapagos, little egrets (subspecies) and island night herons require genetic study. But irrespective of their taxonomic status, they clearly deserve formal conservation planning and action. For these and similar species and populations at risk, species-specific conservation action is needed.
For other herons, conservation most effectively involves preserving and managing their habitats, an effort best enacted through engagement in larger scale habitat, site and multispecies conservation programs. Examples include reserved area management, ecosystem restoration programs, waterbird and wetland management, watershed protection, important area networks, international conventions and flyway initiatives.
The working group model has proven a successful approach. Guidance drawn from the histories of the White-bellied Heron Working Group and Reddish Egret International Working Group might be particularly useful. A yet untried approach is multispecies heron working groups, such as for the great bitterns or Madagascar herons. Such conservation planning takes a series of steps:
recognition, proving of threatened status,
identification of species champions,
holding stakeholder conferences,
establishing formal working groups,
securing institutional support and salaried leaders,
creating range-wide, regional and site-based conservation action plans,
conducting needed research, inventory and monitoring programs,
conducting population size and trend analyses,
identifying risk factors,
securing overhead and project funding,
continual nurturing the working groups,
building capacity for conservation action across the species range,
enacting needed conservation actions,
adaptively revising plans
Beyond species and site conservation planning and execution, global and regional analyses, syntheses, planning, prioritization and facilitation remain necessary. The global leadership of HeronConservation, the IUCN Heron Specialist Group, has proven useful in this. However, it is the engagement of individual members and partners that makes such large-scale undertakings possible. Fortunately, there are now ways for individuals to contribute directly to international syntheses through electronic communication and online publishing facilitated by the specialist group. It is potentially possible for the current global action plan for herons, species accounts, national analyses and biological observations to be updated through volunteer contributions online through the HeronConservation website.
Herons likely will continue to be of popular, scientific, and conservation interest as they respond to changing environmental conditions. In 2022, the Heron Specialist Group celebrated the fortieth anniversary of its founding. Hopefully the next 40 years will see that common heron species continue to prosper, little known herons become better studied and seriously threatened herons are supported by successful conservation action.
Acknowledgements
I thank Chip Weseloh, Katsutoshi Matsunaga and an anonymous reviewer for their comments on an earlier draft of this paper.
Literature Cited
Acharja, I. P., T. Tobgay, L. Lungten, T. Phuntsho, T. Lhendup and S. Tshering. 2022. Saving the critically endangered White-bellied Heron (Ardea insignis) from extinction: two decades of conservation efforts and the way forward. In Research Summaries from the virtual Herons of Worldwide Conservation Concern Symposium, 2021 (Weseloh, D. V. C., M. C. Green, D. Harebottle and K. Matsunaga, eds.). Journal of Heron Biology and Conservation 7: 6. [online].
Betleja, J. 2023. Identification of potential hybrids between Malagasy Pond Heron (Ardeola idea) and Squacco Heron (A. ralloides) based on photos. Journal of Heron Biology and Conservation 8: 2. [online].
Bida, Y. B., R. George, T. Yomcha and S. Khaling. 2022. Setting up protocols to monitor the Critically Endangered White-bellied Heron, Ardea insignis, with focus on threats to the species in the changing landscapes around Namdapha Tiger Reserve, Arunachal Pradesh, India. In Research Summaries from the virtual Herons of Worldwide Conservation Concern Symposium, 2021 (Weseloh, D. V. C., M. C. Green, D. Harebottle and K. Matsunaga, eds.). Journal of Heron Biology and Conservation 7: 6. [online].
BirdLife International. 2016a. Ardea humbloti. The IUCN Red List of Threatened Species 2016: e.T22697012A93598644. [online]. Accessed 06 December 2024.
BirdLife International. 2016b. Egretta eulophotes. The IUCN Red List of Threatened Species 2016: e.T22696977A93596047. [online]. Accessed 06 December 2024.
BirdLife International. 2016c. Egretta vinaceigula. The IUCN Red List of Threatened Species 2016: e.T22696922A93593178. [online]. Accessed 06 December 2024.
BirdLife International. 2017a. Gorsachius magnificus (amended version of 2016 assessment). The IUCN Red List of Threatened Species 2017: e.T22697232A117359084. [online]. Accessed 06 December 2024.
BirdLife International. 2017b. Zonerodius heliosylus. The IUCN Red List of Threatened Species 2017: e.T22697274A117210228. [online]. Accessed 06 December 2024.
BirdLife International. 2020a. Ardea occidentalis. The IUCN Red List of Threatened Species 2020: e.T181501221A181566064. [online]. Accessed 06 December 2024.
BirdLife International. 2020b. Egretta rufescens. The IUCN Red List of Threatened Species 2020: e.T22696916A154076472. [online]. Accessed 06 December 2024.
BirdLife International. 2020c. Gorsachius goisagi. The IUCN Red List of Threatened Species 2020: e.T22697237A154698841. [online]. Accessed 06 December 2024.
BirdLife International. 2021. Ardeola idae. The IUCN Red List of Threatened Species 2021: e.T22697143A189949320. [online]. Accessed 06 December 2024.
BirdLife International. 2023. Agamia agami. The IUCN Red List of Threatened Species 2023: e.T22697200A171903244. [online]. Accessed 06 December 2024.
Bitterns in Rice Project. n.d. Bitterns in Rice Project. [online]. Accessed 06 December 2024.
Browning, M. R. and J. A. Kushlan. 2022. The Great White Heron is a species. Journal of Heron Biology and Conservation 7: 1. [online].
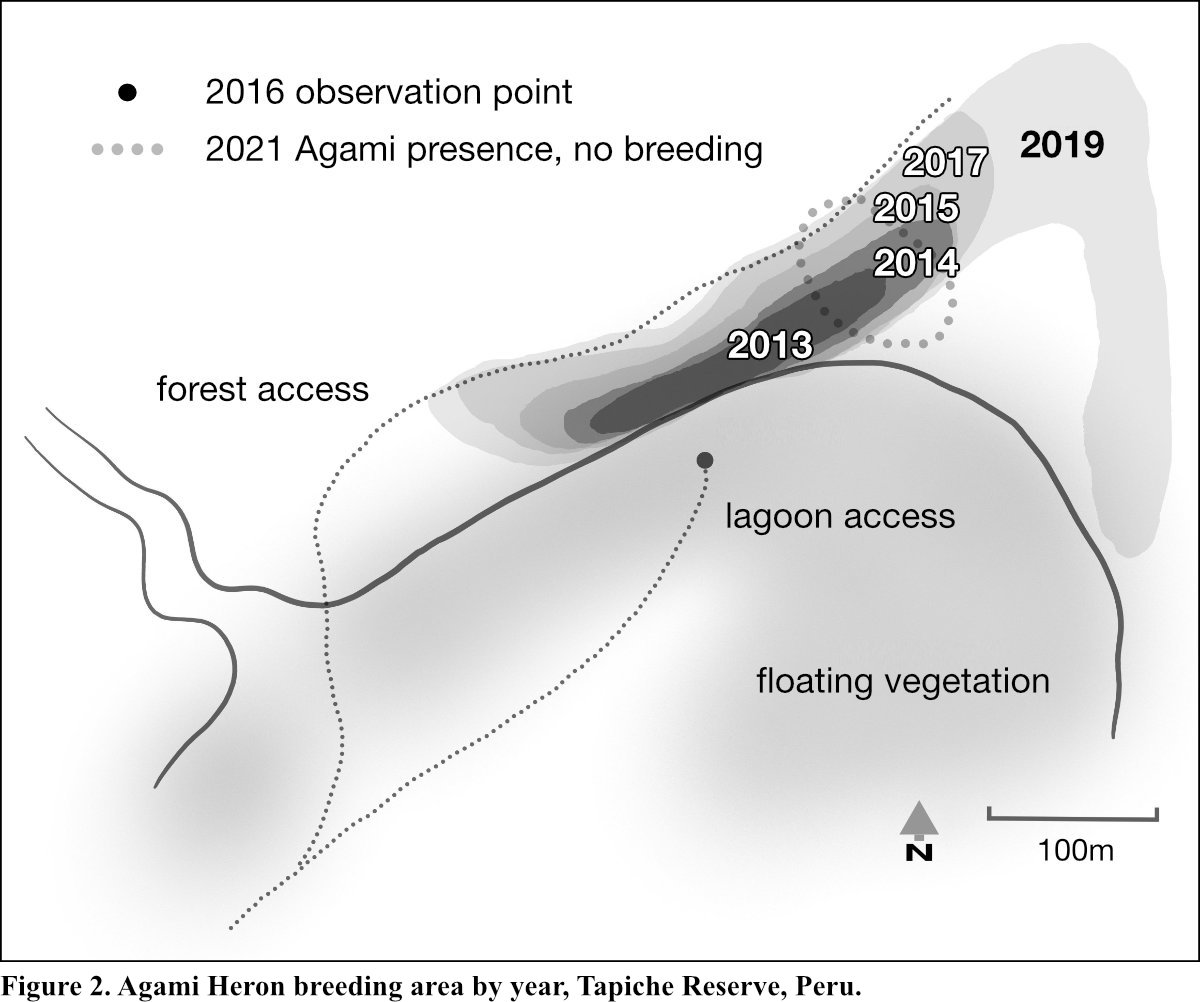
Chen, D., M. D. Reis, J. M. Reintjes and S. O’Donnell. 2022. Observations of the largest known breeding colony of Agami Herons at Tapiche Reserve in the Northeastern Peruvian Amazon. In Research Summaries from the virtual Herons of Worldwide Conservation Concern Symposium, 2021 (Weseloh, D. V. C., M. C. Green, D. Harebottle and K. Matsunaga, eds.). Journal of Heron Biology and Conservation 7: 6. [online].
Cisneros-Heredia, D. and E. Peñaherrera-Romero. 2022. Overview on the diversity, biogeography and conservation of herons in Ecuador. In Research Summaries from the virtual Herons of Worldwide Conservation Concern Symposium, 2021 (Weseloh, D. V. C., M. C. Green, D. Harebottle and K. Matsunaga, eds.). Journal of Heron Biology and Conservation 7: 6. [online].
Collar, N. J., M. J. Crosby and A. J. Stattersfield. 1994. Birds to Watch 2, The World List of Threatened Birds. BirdLife International, Cambridge, U.K.
Commonwealth of Australia 2022. National recovery plan for the Australasian Bittern Botaurus poiciloptilus. [online].
Dine, M. 2012. 2013-2017 Management plan for the White-eared Night Heron (Gorsachius magnificus) at Ba Be National Park, Bac Kan province. People Resources and Conservation Foundation, Hanoi, Vietnam. [online].
Fellowes, J. R., F. Zhou, K. S. Lee, B. C. H. Hau, M. W. N. Lau, V. W. Y. Lam, L. Young and H. Hafner. 2001. Status update on White-eared Night Heron Gorsachius magnificus in South China. Bird Conservation International 11: 101-111.
Fitzsimons, J. A. 2022. Diurnal foraging in short pasture by the endangered Australasian Bittern (Botaurus poieiloptilus) on Kangaroo Island, Australia. Journal of Heron Biology and Conservation 7: 3. [online].
Goodman, G. L. 2022. A coordinated response to the plight of the White-bellied Heron (Ardea insignis). In Research Summaries from the virtual Herons of Worldwide Conservation Concern Symposium, 2021. (Weseloh, D. V. C., M. C. Green, D. Harebottle and K. Matsunaga, eds.). Journal of Heron Biology and Conservation 7: 6. [online].
Green, M. C. 2023. History of the Reddish Egret International Working Group and Conservation Planning. Journal of Heron Biology and Conservation 8: 1. [online].
Green, M. C., D. W. Demarest, B. C. Tarbox, W. A. Cox, J. G. Franco, S. N. Torres, K. L. Stone and W. G. Vermillion. 2022. Conservation Action Plan for the Reddish Egret (Egretta rufescens), 2022 Update. Reddish Egret International Working Group. [online].
Hamaguchi, H., M. Ishikawa, K. Konishi, T. Nagai, H. Oshika and K. Kawakami. 2014. A habitat model for the Japanese Night Heron in the West Mikawa area of Aichi prefecture, Japan. Japanese Journal of Ornithology 63: 33-41.
Hancock, J. A. and J. A. Kushlan. 1984. The herons handbook. Croom Helm, London, U.K.
Hancock, P., M. Muller and A. Flatt. 2006. Report on Slaty Egret breeding in Moremi Game Reserve - April 2006. Babbler 48: 40-43.
He, F., X. Yang, X. Deng, K. Zhu, L. Li and J. Lin. 2011. The White-eared Night Heron (Gorsachius magnificus): from behind the bamboo curtain to the front stage. Chinese Birds 2: 163-166.
He F., F. Zhou, X-J. Yang, L. Yang, X-J. Deng, H-X. Hu, J-S. Lin, H-D. Jiang, Z. Lin, L. Li, K-M. Zhu, B. Anderson, H-H. Zhang and J. T. Dong. 2007. Study on the status of distributions and subpopulations of the White-eared Night Heron Gorsachius magnificus. Acta Zootaxonomica Sinica 32: 802-813.
HeronConservation. 2024. Important Bird Areas for herons. [online]. Accessed 06 December 2024.
HeronConservation. n.d.a. About HeronConservation. [online]. Accessed 06 December 2024.
HeronConservation. n.d.b. Conserving herons: a conservation action plan for herons of the world (Kushlan, J. A. 2007). [online]. Accessed 06 December 2024.
Herring, M. W. 2022. Targeted water management is key to recovery of the Australasian Bittern, Botaurus poiciloptilus. In Research Summaries from the virtual Herons of Worldwide Conservation Concern Symposium, 2021 (Weseloh, D. V. C., M. C. Green, D. Harebottle and K. Matsunaga, eds.). Journal of Heron Biology and Conservation 7: 6. [online].
Herring, M. W., W. Robinson, K. K. Zander and S. T. Garnett. 2019. Rice fields support the global stronghold for an endangered waterbird. Agriculture, Ecosystems and Environment 284: 106599. [online].
Herring, M. W., W. Robinson, K. K. Zander and S. T. Garnett. 2021. Increasing water-use efficiency in rice fields threatens an endangered waterbird. Agriculture Ecosystems and Environment 322: 107638. [online].
Hruska, J. P., J. Holmes, C. Oliveros, S. Shakya, P. Lavretsky, K. G. McCracken, F. H. Sheldon and R, G. Moyle. 2023.. Ultraconserved elements resolve the phylogeny and corroborate patterns of molecular rate variation in herons (Aves: Ardeidae). Ornithology 140: ukad005. [online].
International Union for Conservation of Nature (IUCN). 2024. IUCN Red List of Threatened Species. Version 2024-2. [online]. Accessed 06 December 2024.
Khandu, P. 2022. Nest predation of the critically endangered White-bellied Heron (Ardea insignis) by Masked Palm Civet (Paguma lorvata) in Burichhu, Bhutan. Journal of Heron Biology and Conservation 7: 4. [online].
Kushlan, J. A. 2007. Conserving herons: a conservation action plan for herons of the world. Station Biologique de la Tour du Valat, Arles, France. [online].
Kushlan, J. A. 2012. A history of conserving colonial waterbirds in the United States. Waterbirds 35: 608-625.
Kushlan, J. A. and H. Hafner (eds.). 2000. Heron conservation, Academic Press, London, U.K.
Kushlan, J. A. and J. A. Hancock. 2005. The herons. Oxford University Press, Oxford, U.K.
Kushlan, J. A. and K. N. Hines. 2016. Behavior of the Agami Heron. Waterbirds 39: 187-192.
Kushlan, J. A., M. J. Steinkamp, K. C. Parsons, J. Capp, M. Acosta Cruz, M. Coulter, I. Davidson, L. Dickson, N. Edelson, R. Elliot, R. M. Erwin, S. Hatch, S. Kress, R. Milko, S. Miller, K. Mills, R. Paul, R. Phillips, J. E. Saliva, B. Sydeman, J. Trapp, J. Wheeler and K. Wohl. 2002. Waterbird Conservation for the Americas. Waterbird Conservation for the Americas and US Fish and Wildlife Service, Washington, D.C., U.S.A.
Li, D. 2006. Survey of the status of Nordmann’s Greenshank Tringa guttifer and Chinese Egret Egretta eulophotes in Malaysia. BirdingASIA: 8-9.
McGuire, H. L., S. S. Taylor and F. H. Sheldon. 2019. Evaluating the taxonomic status of the Great White Heron (Ardea herodias occidentalis) using morphological, behavioral and genetic evidence. Auk 136: 1-18.
Ndang’ang’a, P. K. and E. Sande. 2008. International single species action plan for the conservation of the Madagascar Pond-heron Ardeola idae. CMS Technical Series No. 20, AEWA Technical Series No. 39. Bonn, Germany. [online].
New South Wales Government. 2022. Bringing back the bunyip birds - Australian Bittern Conservation Summit 2022. [online]. Accessed 06 December 2024.
Oh, H., Y. Kim and N. Kim. 2010. First breeding record of Japanese Night Heron Gorsachius goisagi in Korea. Ornithological Science 9: 131-134.
Patgiri, S. 2022. Observations and conservation implications of a newly discovered White-bellied Heron Ardea insignis nesting site in India. In Research Summaries from the virtual Herons of Worldwide Conservation Concern Symposium, 2021 (Weseloh, D. V. C., M. C. Green, D. Harebottle and K. Matsunaga, eds.). Journal of Heron Biology and Conservation 7: 6. [online].
Pilgrim, J. D., D. F. Walsh, T. Thanh Tu, N. Duc Tu, J. C. Eames and L. Manh Hung. 2009. The endangered White-eared Night Heron Gorsachius magnificus in Vietnam: status, distribution, ecology and threats. Forktail. Oriental Bird Club 25: 142-146.
Powell, G. V. N. 1983. Food availability and reproduction by Great White Herons, Ardea herodias: A food addition study. Colonial Waterbirds 6: 139-147.
Pratt, T. K. and B. M. Beehler. 2015. Birds of New Guinea. Princeton University Press, Princeton New Jersey, U.S.A.
Price, M. R. S. and G. L. Goodman. 2015. White-bellied Heron (Ardea insignis) conservation strategy. IUCN Species Survival Commission White-bellied Heron Working Group, part of the IUCN SSC Heron Specialist Group. [online].
Pruvot, Y. Z. M. and L. A. Rene de Roland. 2021. Food habits of the Malagasy Pond Heron (Ardeola idae) during the breeding season in northern Madagascar. Journal of Heron Biology and Conservation 6: 1. [online].
Pruvot, Y. Z. M., L. A. Rene de Roland, M. Rakotondratsima, Y. Razafindrakoto, F. Razafindrajao, R. Rabarisoa and R. Thorstrom. 2020. Breeding ecology and nestling growth of the Madagascar Pond Heron (Ardeola idae) in a monospecific colony at Sofia Lake, northern Madagascar. Ostrich 91: 1-13.
Rabarisoa, R. 2022. Current status of the Humblot’s Heron (Ardea humbloti) in Madagascar. In Research Summaries from the virtual Herons of Worldwide Conservation Concern Symposium, 2021 (Weseloh, D. V. C., M. C. Green, D. Harebottle and K. Matsunaga, eds.). Journal of Heron Biology and Conservation 7: 6. [online].
Rabarisoa, R. and R. Hajanirina. in review. Habitat requirements of the Madagascar Pond Heron Ardeola idae in Madagascar. Journal of Heron Biology and Conservation.
Rabarisoa, R., J. Ramanampamonjy, F. Razafindrajao, L-A. R. De Roland and F. Jeanne. 2020. Status assessment and population trends of the Madagascar Pond-Heron (Ardeola idae) from 1993-2016. Waterbirds 43: 45-54.
Reynaud, P. A. and J. A. Kushlan. 2004. Nesting of the Agami Heron. Waterbirds 27: 308-311.
Royal Society for Protection of Nature. 2019. White-bellied Heron Strategic Plan 2019-2039. Royal Society for Protection of Nature, Thimphu, Bhutan.
Royal Society for Protection of Nature. 2022. The White-bellied Heron Conservation Center. Thimphu, Bhutan. [online].
Royal Society for Protection of Nature and Department of Forests and Park Services. 2022. White-bellied Heron Conservation Action Plan (2022-2031). Royal Society for Protection of Nature and Department of Forests and Park Services, Ministry of Agriculture and Forests, Thimphu, Bhutan. [online].
Shafli, S. 2018. Extraordinary discovery in Bihar, India: an apparent White-eared Night Heron Gorsachius magnificus in Valmiki Tiger Reserve. BirdingASIA 29: 9-10.
Stier, A. and J. A. Kushlan. 2015. Agami Heron (Agamia agami) conservation plan. GEPOG Association, Cayenne, French Guiana. [online].
Stier, A., A. Ricardou, S. Uriot, N. de Pracontal and J. A. Kushlan. 2017. Breeding season, home range and migration of the Agami Heron (Agamia agami). Waterbirds 40: 289-296.
Tarbox, B. C., W. A. Cox, J. Franco, K. Stone, W. G. Vermillion and M. C. Green (eds.). 2020. Business plan for conservation of the Reddish Egret in the United States, December 2020. Texas State University, San Marcos Texas, U.S.A. [online].
Tyler, S. 2005. The Slaty Egret Egretta vinaceigula - A review, with special reference to Botswana. Babbler 46: 8-17.
Tyler, S. J. 2012. International single species action plan for the conservation of the Slaty Egret (Egretta vinaceigula). AEWA Technical series No. 43. Bonn, Germany. [online].
Tyler, S. J. 2024. The Slaty Egret Egretta inaceigula - A review, with special reference to Botswana. Journal of Heron Biology and Conservation 9: 1. [online].
White G., J. Purps and S. Alsbury. 2006. The bittern in Europe: A Guide to Species and Habitat Management. The RSPB, Sandy, U.K. [online].
Wilson, T. E., J. Wheeler, M. C. Green and E. Palacios (eds.). 2014. Reddish Egret Conservation Action Plan. Reddish Egret Conservation Planning Workshop, October 2012. Corpus Christi, Texas, U.S.A. [online].
Yanosky, A., R. Irala and T. Galluppi. 2023. Citizen science updates on the occurrence and conservation of herons in Paraguay. Journal of Heron Biology and Conservation 8: 3. [online].
Research Summaries from the virtual Herons of Worldwide Conservation Concern Symposium, 2021
D. V. Chip WeselohCanadian Wildlife Service, 4905 Dufferin St., Toronto, Ontario, M3H 5T4, Canada;
*,
M. Clay GreenDepartment of Biology, Texas State University, 601 University Drive, San Marcus, Texas, 78666, U.S.A.
,
Doug HarebottleRisk and Vulnerability Science Centre, School of Natural and Applied Sciences, Sol Plaatje University, Private Bag X5008, Kimberley, 8300, South Africa
and
Katsutoshi MatsunagaHokkaido Grey Heron Research Group, Hachiken 8 Higashi 5, 4-8-103, Sapporo, Hokkaido, Japan
* Primary contact
Introduction
The second in a series of symposia on Herons of the World was held as part of the 45th Anniversary Meeting of the Waterbird Society during 8-12 November 2021; the entire meeting was held virtually due to the outbreak of Covid-19. This second symposium was originally planned as part of the 15th Pan-African Ornithological Congress (PAOC-15) in Victoria Falls, Zimbabwe, in 2020 but it was postponed, repeatedly, due to Covid-19.
With the postponement of that Congress and Symposium, the Heron Specialist Group (HSG) felt it was necessary to organize a virtual Heron-event that would keep the momentum going from the planning for the African meeting. With at least three heron species of conservation concern in Africa and the many developments with the White-bellied Heron in Bhutan and India, we felt a symposium focused on Herons of Worldwide Conservation Concern would be appropriate, especially in a virtual format where researchers from around the world could contribute without leaving the confines of their home or place of research. Focusing on this group of herons would be an excellent way to draw attention to our most vulnerable species, highlight their plight and showcase the work being done on them as well as make a worldwide appeal for support for their work.
The IUCN (2022), identifies 13 heron species, worldwide, that are of conservation concern. The White-bellied Heron (Ardea insignis) is Critically Endangered and the most threatened heron species in the world. The Endangered species include: the Australasian Bittern (Botaurus poiciloptilus), the White-eared Night-heron (Oroanassa magnifica), the Madagascar Pond-heron (Ardeola idae), the Madagascar Heron (Ardea humbloti) and the Great White Heron (Ardea occidentalis). The Japanese Night-heron (Gorsachius goisagi), the Slaty Egret (Egretta vinaceigula), the Chinese Egret (Egretta eulophotes) and the Agami Heron (Agamia agami) are classified as Vulnerable, and the near Threatened species include: the Reddish Egret (Egretta rufescens), the Forest Bittern (Zonerodius heliosylus) and the Zigzag Heron (Zebrilus undulatus).
We tried to locate researchers for each of these species; we were only partially successful, finding researchers for 7 of the 13 species: the White-bellied Heron, Australasian Bittern, Madagascar Heron and Pond-heron, Slaty Egret, Agami Heron and Reddish Egret. These contributions originated from India-Bhutan (5), Africa, and South and North America (8) and one from Australia. We were unable to locate researchers for: the White-eared Night-heron, Japanese Night-heron, Chinese Egret, Forest Bittern and the Zigzag Heron. We are excited to note that five of the presentations reported on advances in the conservation of the White-bellied Heron, our most endangered heron.
As with our reporting on the first Herons of the World Symposium, we invited the presenters to submit full length papers of their work to either Waterbirds (the journal of the Waterbird Society) or to the HSG’s own journal, the Journal of Heron Biology and Conservation (JHBC). For those who preferred a less onerous format, we invited them to submit an extended abstract of their work for publication in JHBC. The goal of this exercise, as before, was to present a formal update, in the public domain, on the very important conservation work on these selected species.
Below are the extended abstracts from eight of the papers given at the symposium. One full length paper has already appeared in JHBC, four more full length papers are near final and a wrap-up paper (J. Kushlan) is in hand waiting for the outstanding papers to appear. Coincidently, a paper, on the Great White Heron appeared earlier in Volume 7. It was not part of the Symposium but its subject matter is directly relevant. These abstracts and full-length papers provide an important update on the status of more than half of the Herons of Worldwide Conservation Concern.
We extend our greatest appreciation to the researchers who contributed their work on these species of Herons of Worldwide Conservation Concern.
India-Bhutan
Saving the critically endangered White-bellied Heron (Ardea insignis) from extinction: two decades of conservation efforts and the way forward
Indra P. AcharjaRoyal Society for Protection of Nature, Lhado Lam, Thimphu, Bhutan
*,
Tshering TobgayRoyal Society for Protection of Nature, Lhado Lam, Thimphu, Bhutan
,
Lungten LungtenRoyal Society for Protection of Nature, Lhado Lam, Thimphu, Bhutan
,
Thinley PhuntshoRoyal Society for Protection of Nature, Lhado Lam, Thimphu, Bhutan
,
Tshewang LhendupRoyal Society for Protection of Nature, Lhado Lam, Thimphu, Bhutan
and
Sonam TsheringRoyal Society for Protection of Nature, Lhado Lam, Thimphu, Bhutan
* Primary contact
The White-bellied Heron (Ardea insignis) is Critically Endangered and one of the rarest heron species in the world. Fewer than 60 individuals are confirmed to exist today, spanned over Bhutan, India, Myanmar and China within the extent of 165,000 km2 of the Himalayan freshwater ecosystems. However, active nests and breeding populations are only known from Bhutan and India. Since 2015, after the preparation of the conservation strategy and hosting of the first international conference for the species, range countries have been putting efforts to protect the fragmented populations and restore their habitats. In Bhutan, conservation started in 2003 and soon after the first active nest was discovered. It was the first active nest for the country and rediscovery for the world after more than seven decades of the previous record. Over the last two decades, we have monitored the population trend, distribution and habitat use, nest, and active breeding population and mapped major threats to the bird and their habitats. We conducted the annual population survey for the last 19 consecutive years and recorded an average of 23.7 ± 4.4 SD (n = 19) individuals/year. The average annual number of active nests (i.e., number of breeding pairs) found was 2.6 ± 1.4 SD per year (n = 50) with an average clutch size of 2.7 ± 1.4 SD (n = 28). The average hatching success of 1.9 ± 1.1 SD (n = 40) per nest and the average fledging success was 1.8 ± 1.1 SD (n = 42) young per nest. While we observed a nest success rate of 86% (n = 50), there are no indications of population growth. The global population size is on a declining trend and the distribution range is shrinking. Very little information is available on predators, post-fledging survival, dispersal and mortality. Within Bhutan, despite good breeding success, the population has remained low and potentially declining. However, we have no data on yearling/juvenile survival rate. In the recent years, their distribution has shifted from altitudes between 600-1,500 m to wider regions between 100-1,800 m, undoubtedly as a consequence of the activities of humans (rafting, hydro projects and general disturbance). There is also a noticeable decrease in population in some of the older habitats and most of the oldest nesting sites have been abandoned. The small and fragmented population with a restricted range and small gene pool is further threatened by habitat loss due to infrastructure development, hydropower dams, extractive industries, and climate change in the region. Our long-term monitoring and conservation has filled many information gaps and provides important implications including the need for securing ex-situ gene pool, conservation breeding, more coordinated and impactful in-situ conservation efforts to save this species from extinction.
Monitoring the Critically Endangered White-bellied Heron, Ardea insignis, in Namdapha Tiger Reserve, Arunachal Pradesh, India
Yumlam Benjamin BidaAshoka Trust for Research in Ecology and the Environment, Regional Office Eastern Himalaya-Northeast India, NH 10 Tadong, Gangtok, Sikkim, India
,
Rohit GeorgeAshoka Trust for Research in Ecology and the Environment, Regional Office Eastern Himalaya-Northeast India, NH 10 Tadong, Gangtok, Sikkim, India
,
Tajum YomchaNamdapha Tiger Reserve, Miao, Changlang District, Arunachal Pradesh, India
and
Sarala KhalingAshoka Trust for Research in Ecology and the Environment, Regional Office Eastern Himalaya-Northeast India, NH 10 Tadong, Gangtok, Sikkim, India
*
* Primary contact
The White-bellied Heron (WBH, Ardea insignis) is one of the most threatened birds in the world. India may host the largest population of WBH, but there have been limited population surveys. Namdapha Tiger Reserve (NTR), in the north-east of the Indian state of Arunachal Pradesh, has been noted as a strong-hold for the species where sightings have been recorded and estimates thus far are of 5-6 individuals remaining. However, regular monitoring of the species has been a challenge due to lack of resources and with more attention being given to monitoring large and charismatic mammals found in the Tiger Reserve. The purpose of this study was to assess the presence of the WBH in this region.
This study was carried out during 2021-2022. We laid out 32 transects each 1-2 km in length and 30-50 m in width in the known and potential habitats of WBH along the Deban, Noa-dehing and Namdapha rivers in NTR. These transects were walked (by 2-3 observers) searching for the WBH in the period between October 2021-March 2022. Each transect was monitored four times. Environmental parameters, activities of the birds and habitat characteristics were recorded. These preliminary surveys resulted in 18 direct sightings of the WBH including at least ten different birds. Eight of these sightings were repeats specifically in transects which were located in sites that were previously reported to have WBH in NTR. Six of the direct sightings were in sites from where WBH has never been sighted or reported in published literature before.
Anthropogenic activities like fishing, using river banks as thoroughfare, road construction, agriculture activities and sheer presence of a large number of people in pristine WBH habitats have the potential to be a direct threat to the species or its habitat. We hope some of these are short term and will reduce drastically when the Miao-Vijanaygar road (which cuts across the core area of the Reserve) construction is completed. While areas in core areas of NTR may provide a better habitat for the species in terms of lesser threats and disturbances, it is imperative to monitor the species in its known habitats as establishing its presence in unknown areas will be challenging considering the terrain and the shy and elusive nature of the bird.
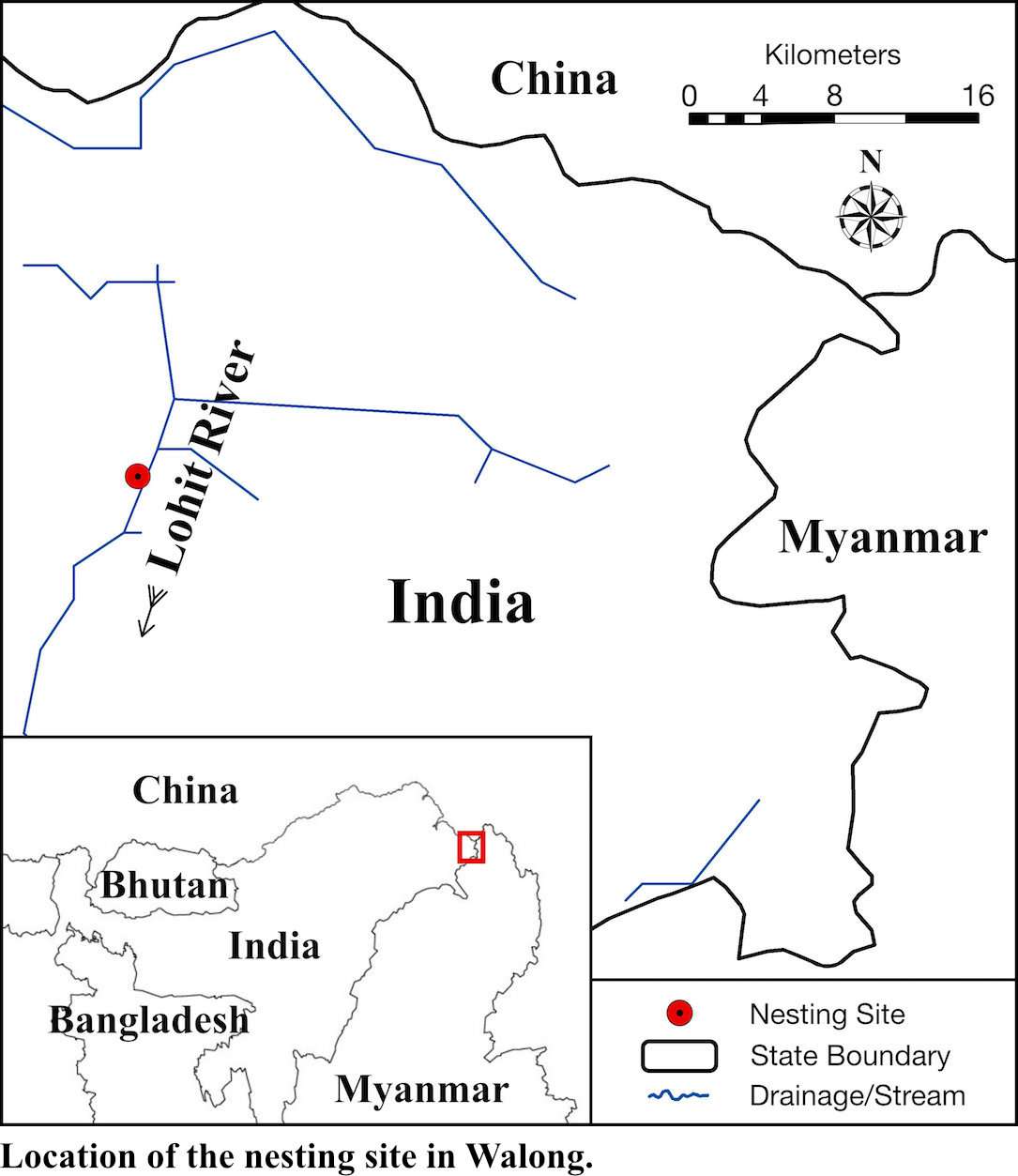
Implications and observations of a newly discovered White-bellied Heron Ardea insignis nesting site in India
Samiran PatgiriDepartment of Forestry, Mizoram University, Aizawl, Mizoram, India
The White-bellied Heron Ardea insignis is a Critically Endangered bird with an estimated global population of around 200-250 individuals. According to the IUCN SSC White-bellied Heron Working China Group, the confirmed population throughout its range is less than 60 individuals. The White-bellied Heron is presently distributed in Bhutan, Northeast India and Myanmar. On 16 April 2021, a pair of White-bellied Heron was recorded along with a nest in Walong, adjacent to the Indo-border in Arunachal Pradesh. The nest was recorded in a Merkusi’s Pine (Pinus merkusii) forest close to Walong Township at 1,250 m asl on river Lohit, about 11 m from the ground. This is the second nesting record in India. Previously Namdapha Tiger Reserve was the only nesting site in India with a population of at least 7-8 individuals. The record in Walong is vital due to the proximity of Walong to other White-bellied Heron habitats like the Namdapha and Kamlang Tiger Reserves. It is also close to the priority sites which the Working Group identified. The sightings in Walong give us the scope to explore other areas close to Walong, including China. Preliminary surveys were carried out in Walong and the nearby areas to understand their habitat, behaviour, the associated threats, etc. Our observations show that the birds are highly adapted and tolerant toward humans. It is also significant since the birds were nesting and occupying the areas very close to human settlements, which contradicts our established perception that the White-bellied Heron in India is shy and avoids human presence. Also, household questionnaire surveys were carried out in 14 villages, including the Walong Township and the villages covering the areas from Kaho (the nearest village to the Indo-China border) to villages downstream from Walong Township. In the Kobo Collect mobile app, a semi-structured questionnaire was created. The questionnaire surveys were mainly directed toward men since they are primarily engaged in hunting. Sixty-four people responded to the questionnaires. Our findings show that 59% of the respondents had never seen a White-bellied Heron, 12% had seen it recently, and 29% had seen it in recent years. This pattern indicates that the species could have mainly remained unnoticed due to their small number and camouflaged nature. We could not find any hunting records of the birds, but the Meyor and the Miju Mishimis, the dominant tribes of the area, practice traditional hunting. We also found traces of forest fire in the nesting site.
Location of the nesting site in Walong.
Landscape of Lohit River.
A Coordinated response to the plight of the White-bellied Heron (Ardea insignis)
The Critically Endangered White-bellied Heron (Ardea insignis) is the one of the world's rarest birds and is the second largest heron. As such, a group of international and range-state based conservationists, government representatives and scientists, came together in Guwahati, India, in December 2014 for a Conservation Strategy Workshop. As a result, the White-bellied Heron Working Group was formed under the IUCN SSC Heron Specialist Group; International and Regional Coordinators were appointed and a global Conservation Strategy (Price and Goodman 2015), was produced in November 2015. This was followed by a second International Workshop held in Bhutan in late November 2015. Survey work in China got underway in 2015 to try and understand the likelihood that a population still survived in China or not, sadly no birds were located. However, in December 2019, a young bird was found injured and rapid action on the ground saw the bird enter captivity to later be released. The collective and expeditated effort for this single individual was impressive – involving scientists and zoo professionals from China, Japan, Singapore and Europe. Sadly, the bird did not survive, and its satellite transmitter was later found having been deliberately removed by people.
In India, the White-bellied Heron Coordinator held national meetings and field work increased subsequent to the two international workshops. The Working Group was able to locate some funding for parts of this field work and in 2018 help to organise a workshop and publish a subsequent report on priority sites for survey within India (White-bellied Heron Working Group 2019). That publication has since been guiding field work being undertaken by various individuals and organisations and in-fact identifying new records of birds in India.
Soon after the second international workshop, one of the top priority actions was implemented when two birds were fitted with satellite transmitters in Bhutan in June 2016. Unfortunately, the data were poor but the training delivered by a European transmitter expert to the team in Bhutan was invaluable. In 2020, the captive breeding centre for White-bellied Heron was finally completed by the Royal Society for Protection of Nature (RSPN) and the Bhutanese government and despite the presence of COVID, three fledglings were brought into captivity in 2021. The Bhutanese team had undergone two previous training efforts (at European zoos to work with large waterbirds which might have similar needs and risks) to help prepare for the role of caring for one of the world’s most threatened birds. This was organised via the Working Group (starting in 2017) and appears to have helped the capable team at RSPN. In recent months, an injured bird has been found and also brought into the breeding centre. The plans continue to harvest chicks and/or eggs from the wild in subsequent years and to increase the capacity of the centre as soon as possible.
The IUCN SSC White-bellied Heron Working Group has been working through its members to provide multi-pronged support at levels ranging from funding to capacity building. There is now a small yet committed community of implementors and advisors placed across range states and the globe. While the role to coordinate this group could be more fully realised, having any such working group in the context of such an underfunded and overlooked species has proven that traction can be had with more concerted and coordinated efforts. In the future, it is hoped that India and Bhutan will remain the strongholds of the species and that protection can continue and be scaled up, while captive breeding efforts can come to fruition to provide a security stock for the species.
Literature Cited
Price, M. R. S. and G. L. Goodman. 2015. White-bellied Heron (Ardea insignis): Conservation strategy. IUCN Species Survival Commission White-bellied Heron Working Group, part of the IUNC SSC Heron Specialist Group. [online]
White-bellied Heron Working Group. 2019. Prioritising search areas for White-bellied Heron in India. IUCN Species Survival Commission White-bellied Heron Working Group, part of the IUNC SSC Heron Specialist Group. [online]
Australasian
Targeted water management is key to recovery of the endangered Australasian Bittern, Botaurus poiciloptilus
Matthew W. HerringMurray Wildlife, Biodiversity Research, Education and Conservation Consultancy, Australia; www.murraywildlife.com.au
Charles Darwin University, Ellengowan Dr., Casuarina NT 0810, Australia;
The Australasian Bittern (Botaurus poiciloptilus) is a cryptic, globally endangered waterbird and the most threatened bittern in the world. Over the past decade, concerted efforts, including substantial public funding, have promoted the species as a flagship for wetland conservation and provided new insights into the conservation status, key threats and required conservation actions. The global population appears to be around 2,000 individuals and still in decline, particularly in New Zealand. The new Action Plan for Australian Birds estimates the national population at 1,300 birds (Herring et al. 2021a), concentrated in the Riverina region of New South Wales, where rice fields support 500-1,000 birds, with the most important natural wetlands comprising the Barmah-Millewa, Lowbidgee and Fivebough-Tuckerbil systems. Northern Victoria, especially around the Kerang region, contains several key wetlands, e.g., Hird and Johnson Swamp, while southwestern Victoria, including adjacent parts of southeastern South Australia, can also support relatively large numbers, notably at Bool and Hacks Lagoon, and Pick Swamp. With the exception of rice fields, almost all of these bitterns occur in protected areas and game reserves. Tasmania and southwestern Australia each support a small, relatively isolated subpopulation of less than 100 individuals. In Australia, the increasing severity and frequency of drought is now considered the key threat, emphasizing the impacts of climate change and the importance of drought refuges. Dry periods reduce the environmental water available for key bittern sites in the Murray-Darling Basin and amplify water-use efficiency measures in rice fields that are undermining successful breeding opportunities (Herring et al. 2021b). Improved water management across all wetland types could maximize the benefits to bitterns. For example, providing a sufficient hydroperiod for successful breeding that also incorporates a drying phase can maintain the preferred early successional stages of vegetation and maximize prey abundance. Incentives for bittern-friendly rice farming and targeted environmental water management at key wetlands should be prioritized, while the potential impact of fox and cat predation needs to be assessed. Despite increased attention, the conservation status of the Australasian Bittern remains grave and greater management effort is urgently required.
Literature Cited
Herring, M. W., P. Barratt, A. H. Burbidge, M. Carey, A. Clarke, S. Comer, B. Green, R. Pickering, C. Purnell, A. Silcocks, V. Stokes, E. Znidersic, R. P. Jaensch and S. T. Garnett. 2021a. Australasian Bittern Botaurus poiciloptilus. Pages 222-224 in The Action Plan for Australian Birds 2020 (S. T. Garnett and G. B. Baker, eds.). CSIRO Publishing, Melbourne, Australia.
Herring, M. W., W. Robinson, K. K. Zander and S. T. Garnett. 2021b. Increasing water-use efficiency in rice fields threatens an endangered waterbird. Agriculture, Ecosystems and Environment 322: 107638. [online]
Africa
Current status of the Humblot's Heron (Ardea humbloti) in Madagascar
Rivo RabarisoaAsity Madagascar, BP 1074, Antananarivo 101, Madagascar;
The Endangered Humblot's Heron (also known as Madagascar Heron, Ardea humbloti) breeds in Madagascar with a recent record in Mayotte. To strengthen its conservation, investigations were carried out through literature reviews and field expeditions undertaken from 1993 to 2019 in Madagascar for population assessment and trend evaluation. Waterbird censuses have been conducted twice a year when possible. Trend analyses were conducted using Trends and Indices for Monitoring data (TRIM) software based on 26 years of survey and population estimates from 2010 to 2019. Missed values for unvisited sites were evaluated to complete the count by using indices from monitoring data. Based on 683 records, the species occurred in various types of wetlands from marine and coastal habitats to inland wetlands with reconstructed habitats throughout Madagascar. The Humblot's Heron displayed a skewed distribution with higher concentration along the western coastal area, it becomes rare in the southern part and absent along the eastern part of the country. The 2019 population was evaluated at 1,290 mature birds with a minimum of 645 breeding pairs. Most (59.1 %) of the recorded population of the species occurred inside Protected Areas in Madagascar, where several instances of large numbers were noted, e.g., Manambolomaty lakes complex (137 individuals, 2003), Mahavavy Kinkony wetland complex (49 individuals, 2017), Baly Bay (43 individuals, 2000) and Mangoky Ihotry wetland complex (37 individuals, 2019). The species was recorded nesting in colonies but some of the time (31.4 %) it was seen alone, with one record in Mayotte. This heron breeds all year with 76.6% of breeding records seen between September to January. Most of its breeding area was recorded in western Madagascar. The population showed a moderate but significant decline of 1.4% per year (P < 0.01). The main threats are 1) habitat destruction such as slash and burn activities to convert wetlands to agricultural lands, 2) impacting reproductive success through the collection of eggs and fledglings, especially at their breeding colony sites, 3) human disturbance at nesting and foraging sites by over-fishing activities and the collection of aquatic plants, 4) the invasion of alien species, mainly the Water Hyacinth (Eichhornia crassipes), which limits the heron’s foraging area and 5) the possible hybridization with its congener the Grey Heron (Ardea cinerea), of which at least one possible case was seen (see photo). A Humblot's Heron Action Plan is needed to preserve this species including: 1) reinforcing conservation action inside Protected Areas, 2) strengthening mass media communication and education about the species, 3) establishing a Community Conservation Group outside of the Protected Area and 4) completing a bioecological study of the species in Madagascar.
The Humblot's Heron and the Grey Heron together at the same nest. Besalampy, Madagascar, June 2018 (Photo by Rivo Rabarisoa).
Literature Consulted
Jeanne, F., A. Laubin, B. Ousseni, C. Crémades, C. Pusinéri and P. Lizot. 2015. Bilan 2010-2015 des Ardéidés nicheurs et menacés de Mayotte. Gepomay, Mayotte, France.
Langrand, O. 1995. Guide des oiseaux de Madagascar, Delachaux and Niestlé, Lausanne, Switzerland.
Rabarisoa, R. 2001. Variation de la population des oiseaux d’eau dans le complexe des lacs de Manambolomaty, un site Ramsar de Madagascar. Ostrich 72 (Supplement 15): 83-87.
Rabarisoa, R., O. Rakotonomenjanahary and J. Ramanampamonjy. 2006. Waterbirds of Baie de Baly, Madagascar. Pages 374-375 in Waterbirds around the World (G. C. Boere, C. A. Galbraith and D. A. Stroud, eds.). The Stationery Office, Edinburgh, U.K.
Sartain, A. and A. F. A. Hawkins (eds.). 2013. The birds of Africa. Volume VIII: The Malagasy region. Christopher Helm, London, U.K.
South America
Observations of the world’s largest known breeding colony of Agami Herons (Agamia Agami) at Tapiche Reserve in the Northeastern Peruvian Amazon
Deborah ChenTapiche Jungle Reserve, Loreto, Peru;
*,
Murilo D. ReisTapiche Jungle Reserve, Loreto, Peru
,
Jana M. ReintjesTapiche Jungle Reserve, Loreto, Peru
and
Sean O’DonnellDepartment of Biodiversity, Earth and Environmental, Science (BEES) and Biology, Drexel University, Philadelphia, Pennsylvania, U.S.A.
* Primary contact
The Agami Heron (Agamia agami) is widespread in the Neotropics but remains a poorly understood species; little has been confirmed regarding population sizes and trends, seasonal migration patterns and the Agami’s role in the wetland communities it inhabits. In this study, we explore these topics through observations made from 2013 to 2021 of a large mixed-species heron breeding colony, including Agami Herons, located at the Tapiche Reserve in northeastern Peru. Situated in a seasonally flooded forest at the edge of a lagoon, the two-hectare breeding colony is observable via boat. An exhaustive census of the entire breeding area was not possible due to the large size of the colony and inadequate personnel and resources to complete the task. For the March 2017 inventory, we applied the area count technique in transects of approximately 100 m2 spaced at five intervals along the length of the colony. The count was taken when the first Agami hatchlings were about 5-7 days old, and the Agami Heron was the only colonial heron species nesting in this area. Using the formula in the Agami Heron Working Group protocol for extrapolation, (the number of nests in the counted surface × real colony surface/counted surface), the estimated number of Agami nests in 2017 was 15,450. An example of nest density at the nesting site can be seen in Figure 1. The Agami were typically present at this site between January and July, outside of which the colony site was deserted. Occupation of the area is shown by year in shades of grey in Figure 2. Observation was limited to a single access point in 2016 due to an overgrowth of floating vegetation. The Agami were completely absent in 2018, when the nesting area did not flood, and in 2020, when the flood was low and delayed. The Agami Herons were the first herons to occupy the breeding ground during years of breeding success, with Boat-billed Herons (Cochlearius cochlearius) arriving 4-8 weeks later in equal thousands of numbers as the Agami. The Boat-billed Herons typically arrived around the time the earliest Agami hatchlings were old enough to cling to branches outside the nest. The Boat-billed Herons appeared to use existing Agami nests rather than building new structures, and Agami fledglings were often observed perched just next to nests occupied by Boat-billed Herons incubating eggs. In the years that the Agami did not nest, the Boat-billed Herons arrived in far fewer numbers and also failed to breed. Territorial disputes between the Agami and Boat-billed Herons were common. Eight additional waterbird species were resident in the rookery area, some breeding concurrently with the Agami. These included Cocoi Heron (Ardea cocoi) - very common, Neotropic Cormorant (Nannopterum brasilianus) - very common, Anhinga (Anhinga anhinga) - very common, Hoatzin (Opisthocomus hoazin) - very common, Great Egret (Ardea alba) - very common, Black-crowned Night Heron (Nycticorax nycticorax) - common, Snowy Egret (Egretta thula) - common and Wattled Jacana (Jacana jacana) - very common. Nearby, in other parts of the lagoon, were also Striated Heron (Butorides striata) - very common, Horned Screamer (Anhima cornuta) - very common and Azure Gallinule (Porphyrio Flavirostris) - uncommon. Primates, raptors and reptiles were seen preying upon Agami eggs and young. Prior to the establishment of the reserve in 2010, local people exploited the rookery by collecting eggs and hunting adult birds almost to the extinction of the colony. The Agami responded rapidly and positively to protection of the rookery and showed resilience to isolated episodes of drought. Some observations suggest, however, that these birds are challenged by consecutive years of adverse climate conditions. An understanding of the Agami’s foraging behavior and range while breeding, movement patterns outside the breeding season and alternate breeding locations, if any, during atypical flood years would be of immense value to conservation efforts for this iconic heron species.
Figure 1. An example of nest density at the nesting area in April 2017. Nests are marked with pink dots and adult Agami are marked with orange dots.
Figure 2. Agami Heron breeding area by year, Tapiche Reserve, Peru.
Overview on the diversity, biogeography and conservation of Herons in Ecuador
Diego Cisneros-HerediaUniversidad San Francisco De Quito USFQ, Colegio De Ciencias Biológicas Y Ambientales, Instituto IBIOTROP, Laboratorio De Zoología Terrestre & Museo De Zoología, Quito 170901, Ecuador
* and
Emilia Peñaherrera-RomeroUniversidad San Francisco De Quito USFQ, Colegio De Ciencias Biológicas Y Ambientales, Instituto IBIOTROP, Laboratorio De Zoología Terrestre & Museo De Zoología, Quito 170901, Ecuador
* Primary contact
Twenty-three species of herons have been reported in Ecuador, including Agami Heron (Agamia agami) classified under the IUCN threatened category of Vulnerable and Zigzag Heron (Zebrilus undulatus) considered as Near Threatened. Few studies have focused explicitly on the herons of the country, and little information is available about their current distribution, natural history and ecology, population trends and conservation status. We produced an integrated and updated assessment of the species richness and biogeographic patterns for all species and subspecies of herons of Ecuador. This study is based on a large species occurrence dataset obtained from different sources, including fieldwork, scientific literature, grey literature, natural history museums, open data biodiversity databases and private expert databases. Analyses emphasise wetlands and other localities that are part of the National System of Protected Areas of Ecuador or are classified as Important Bird Areas or RAMSAR sites. We identified causes driving population changes (declines or increases), including habitat changes, illegal hunt, invasive species or human-heron conflicts. Finally, we applied the IUCN Red List categories and criteria to evaluate the extinction risk for all species at the national level and provide suggestions for implementing research and conservation actions.
A Conservation Action Plan for the Herons of the World
James A. Kushlan
Table 1. Species and populations of herons showing species conservation status (SCS), distribution,population conservation status (PCS), 1% conservation population threshold, and population estimate.
English Name
Scientific Name
SCS
Population
Range
PCS
1%
Population
Grey Heron
Ardea cinerea
LC
A.c.cinerea
SubSahara Africa
LC
10000
A.c.cinerea
W Europe, NW Africa
LC
2700
290000-340000
A.c.cinerea
C & E Europe
LC
2200
189000-256000
A.c.cinerea
C, SW Asia
LC
A.c.cinerea
S Asia
LC
1000
100000
A.c.jouyi
E Asia
LC
10000
A.c.jouyi
Sumatra
VU
15
1000-2000
A.c.monicae
Mauritania
VU
100
7500-12500
A.c.firasa
Madagascar, Seychelles
EN
50
5000
Great Blue Heron
Ardea herodias
LC
A.h.herodias
N, Central America
LC
124500
A.h.herodias
W North America
LC
(in above)
A.h.herodias
N Mexico
LC
(in above)
A.h.herodias
Baja Mexico
LC
A.h.fannini
Alaska, W Canada
LC
65
6500
A.h.wardi
North America
LC
A.h.occidentalis
Florida, Caribbean
VU
1500
A.h.occidentalis
Caribbean
VU
A.h.occidentalis
Central America
VU
A.h.cognata
Galapagos
LC
Cocoi Heron
Ardea cocoi
LC
South America
LC
10000
White-necked Heron
Ardea pacifica
LC
Australia, Oceania
LC
1000
Great Egret
Ardea alba
LC
A.a.alba
Europe
VU
470
38000-54000
A.a.alba
SW Asia
LC
1000
A.a.melanorhynchos
Tropical Africa
LC
3000
100000-500000
A.a.egretta
North America
LC
2700
270000
A.a.egretta
Mexico
LC
A.a.egretta
Central America
LC
A.a.egretta
Caribbean
LC
A.a.egretta
South America
LC
Eastern Great Egret
Ardea modesta
S Asia (nb)
LC
1000
E Asia (nb)
LC
1000
Indonesia
LC
Australia
LC
1000
New Zealand
VU
1
100
Intermediate Egret
Ardea intermedia
LC
A.i.intermedia
S Asia
LC
1000
A.i.intermedia
E, SE Asia, C Japan, Indonesia
LC
1000
Indonesia
LC
A.i.brachyrhyncha
SubSahara Africa
LC
1000
A.i.plumifera
E Indonesia, New Guinea, Australia
LC
10000
Black-headed Heron
Ardea melanocephala
LC
Africa
LC
3000
100000-500000
Malagasy Heron
Ardea humbloti
EN
Madagascar
EN
20
1000-3000
White-bellied Heron
Ardea insignis
EN
SE Asia
CE
6
250-1000
Sumatran Heron
Ardea sumatrana
LC
SE Asia
LC
1000
Australia
VU
75
7500
Goliath Heron
Ardea goliath
LC
SubSahara Africa
LC
1000
SW Asia
EN
1
50
Purple Heron
Ardea purpurea
LC
A.p.purpurea
Tropical Africa
LC
880
75000-100000
A.p.purpurea
SW, NW Europe, NW Africa
LC
120
11500-12100
A.p.purpurea
C, E Europe, Mediterranean
LC
2200
13500-300000
A.p.purpurea
SW Asia (br)
LC
250
A.p.madagascariensis
Madagascar
VU
75
5000-10000
A.p.bournei
Cape Verde Island
EN
1
<50
A.p.manilensis
S Asia
LC
250
25000
A.p.manilensis
SE, E Asia
LC
1000
Cattle Egret
Ardea ibis
LC
A.i.ibis
S Africa
LC
10000
Tropical Africa
LC
NW Africa
LC
1300
100000-150000
SW Europe
LC
2800
250000-310000
E Mediterranean, SW
LC
1000
Asia
LC
North America (USA, Canada)
LC
750000-1500000
Mexico
LC
Central America
LC
Caribbean
LC
South America
LC
A.i.seychellarum
Seychelles
LC
100
A.i.coromandus
S Asia
LC
A.i.coromandus
E, Se Asia
LC
10000
A.i.coromandus
Australia, New Zealand, New Guinea
LC
10000
100000
Green Heron
Butorides virescens
LC
B. virescens
C, E North America
LC
B. virescens
Mexico
LC
B. virescens
Central America
LC
B. virescens
Caribbean
LC
B. virescens
Bermuda
VU
1
30
B.v.bahamensis
Bahamas
LC
B.v.anthonyi
W North America, N Baja California
LC
B.v.frazari
S Baja California
LC
Striated Heron
Butorides striata
LC
B.s.striata
Central America (Panama), N South America
LC
B.s.striata
C South America
LC
B.s.striata
Bolivia
LC
B.s.sundevalli
Galapagos
LC
B.s.brevipes
E Africa, SW Asia
LC
250
B.s.atricapilla
SubSahara Africa
LC
10000
B.s.rutenbergi
Madagascar
LC
250
B.s.rhizophorae
Comoro Islands
LC
100
B.s.crawfordi
Aldabra, Amirante
LC
50
5000
B.s.degens
Seychelles
LC
100
B.s.albolimbata
Chagos Islands, Maldives, Diego Garcia
VU
100
B.s.chloriceps
India
LC
250
25000
B.s.spodiogaster
W Sumatra, Andaman, Nicobar
LC
B.s.amurensis
NE China, SE Russia, Korea, Japan, Ryukyu, Bonin, Taiwan, S China
LC
B.s.actophila
S China, N Indonesia, N Myanmar
LC
B.s.javanica
W Indonesia, Indian Ocean, Myanmar, Thailand
LC
1000
B.s.moluccara
Moluccas
LC
B.s.moluccara
NW New Guinea, Aru
LC
B.s.solomonensis
New Ireland, Solomon Islands, Vanuatu, W Fiji
LC
250
B.s.idenburgi
W Papua
LC
B.s.flyensis
S New Guinea
LC
B.s.stagnatilis
N Australia
LC
B.s.stagnatilis
NW Australia
LC
B.s.stagnatilis
NE Western Australia
LC
B.s.patruelis
Tahiti, Society
LC
2
100-200
B.s.macrorhyncha
E Australia
LC
B.s.macrorhyncha
SC New Guinea, NE Queensland
LC
B.s.carcinophila
Taiwan, Philippines, Sulawesi
LC
B.s.steini
Lesser Sundras, Indonesia
LC
Squacco Heron
Ardeola ralloides
LC
A.r.ralloides
SW Europe, NW Africa
LC
40
2700-5600
A.r.ralloides
C, E Europe, E Mediterranean
LC
600
24000-76000
A.r.ralloides
SW Asia
LC
1000
A.r.paludivaga
SubSahara Africa, Madagascar
LC
4500
300000-600000
Indian Pond Heron
Ardeola grayii
LC
SW Asia S Asia
LC
10000
Myanmar, Andaman, Nicobar
LC
Maldives
VU
Chinese Pond Heron
Ardeola bacchus
LC
E, SE, S Asia
LC
Javan Pond Heron
Ardeola speciosa
LC
A.s.speciosa
W C Indonesia, S Philippines
LC
A.s.continentalis
C Thailand, S Indochina
LC
1000
Malagasy Pond Heron
Ardeola idae
EN
Madagascar, Aldabra, E Africa
EN
40
2000-6000
Rufous-bellied Heron
Ardeola rufiventris
LC
E & S Africa
LC
1000
Reddish Egret
Egretta rufescens
LC
E.r.rufescens
S USA
VU
250
9000-15000
E.r.rufescens
Mexico
VU
15
1500
E.r.rufescens
Central America
VU
100
E.r.rufescens
Caribbean
VU
100
E.r.rufescens
N South America
VU
275
≤>2250-3000
E.r.dickeyi
San Luis Island, W Mexico
VU
Pied Heron
Egretta picata
LC
Oceania
LC
1000
Slaty Egret
Egretta vinaceigula
VU
C Africa
VU
40
3000-5000
Black Heron
Egretta ardesiaca
LC
SubSahara Africa
LC
1000
Madagascar
NT
Tricolored Heron
Egretta tricolor
LC
E.t.tricolor
N South America
LC
≤>180000
E.t.ruficollis
USA
LC
293000
E.t.ruficollis
Mexico
LC
>1500
E.t.ruficollis
Central America
LC
E.t.ruficollis
Caribbean
LC
White-faced Heron
Egretta novaehollandiae
LC
E.n.novaehollandiae
New Zealand, Australia, New Guinea, E Indonesia
LC
>5000
E.n. parryi
NW Australia
LC
Little Blue Heron
Egretta caerulea
LC
N America
LC
2600
225000-300000
Mexico
LC
1100
75000-150000
Central America
LC
Caribbean
LC
South America
LC
>150000-285000
Snowy Egret
Egretta thula
LC
E.t.thula
Central and North America
LC
2100
215000
E.t.brewsteri
W North America, Baja California
LC
10000
E.t.thula
Mexico
LC
1100
75000-150000
E.t.thula
Central America
LC
E.t.thula
Caribbean
LC
E.t.thula
South America
LC
>390000-1170000
Little Egret
Egretta garzetta
LC
E.g.garzetta
SubSahara Africa
LC
3500
200000-500000
E.g.garzetta
W Europe, NW Africa
LC
1300
200000-500000
E.g.garzetta
C, E Europe
LC
580
44000-72400
E.g.garzetta
Barbados
VU
1
30-60
E.g.garzetta
W Asia
LC
1000
E.g.garzetta
S Asia
LC
1500
100000-200000
E.g.garzetta
E SE Asia
LC
10000
E.g.nigripes
Java, New Guinea
LC
E.g.immaculata
Australia, New Zealand
LC
1000
E.g.gularis
W Africa
LC
1000
E.g.schistacea
NE Africa, Red Sea
LC
1000
E.g.schistacea
SW Asia
LC
170
17000
E.g.dimorpha
Madagascar
LC
130
6000-20000
E.g.dimorpha
Aldabra, Amirante
LC
60
3000-9000
E.g.dimorpha
E Africa
LC
100
10000
Chinese Egret
Egretta eulophotes
VU
E Asia
VU
30
2600-3400
Eastern Reef Heron
Egretta sacra
LC
E.s.sacra
SE Asia, Andaman, Nicobar, China, Taiwan, Korea, Japan, Indonesia, Philippines
A Conservation Action Plan for the Herons of the World
James A. Kushlan
Kushlan, J. A. 2007. Conserving Herons, A Conservation Action Plan for the Herons of the World. Heron Specialist Group and Station Biologique de la Tour du Valat, Arles, France.
Readers are invited to use and reproduce the text, provided that the report is fully cited and credit is given to the Heron Specialist Group and to the Tour du Valat. All photo rights are reserved by the publisher, from whom written permission is required for use.
Conserving Heronsis dedicated to the memory ofHeinz Hafner and of James Hancock.
Executive Summary
Conserving Herons is the conservation action plan for the herons of the world. It is a product of the Heron Specialist Group and is the third version of such a planning tool, appearing ten years after the first. The Plan sets forth strategic principles for heron conservation, which focus on conservation of populations, habitat, sites, and flyways outlining specific global and regional conservation projects.
This edition of the plan assesses the conservation status of 62 extant species and 259 populations. Assessing conservation status of species is a cooperative venture among the Heron Specialist Group, the waterbird population estimates program of the Ramsar Convention Bureau and Wetlands International and the species conservation assessment program of the World Conservation Union (IUCN) and BirdLife International. Nine species and 12% of heron populations are assessed as being under threat. Four recent populations are thought now to be extinct. Overall, habitat degradation is the most important threat to herons, and the plan identifies the characteristic habitat for herons of conservation interest. The Plan recognizes the criticality of protecting important sites for herons and endorses sites also identified as Wetlands of International Importance by the Ramsar Bureau and as Important Bird Areas by BirdLife International. As most heron species migrate or otherwise move seasonally, some inter-continentally, the plan identifies flyway conservation as being an important tool in heron conservation.
Partnerships are the principal means of moving heron conservation forward. In addition to the planning and assessment partnerships noted above, the Plan recognizes that heron conservation often can best proceed as part of more-encompassing conservation programs, such as flyway and migratory species initiatives, wetland and coastal conservation programs, continental, regional and national bird conservation initiatives, protected areas conservation, biodiversity conservation, and local conservation actions. The latter are the most critical, as heron conservation ultimately depends on site and habitat conservation, which are primarily local matters. The sustainable use of habitats by both people and herons managed at the local level is considered to be the ultimate key to heron conservation.
Herons are overall an adaptable group of birds, most of which have been able to co-exist with humans over much of their natural ranges. Most heron populations are not facing immediate threat. Those that are at conservation risk, for the most part, are facing crises in habitat, which can be addressed through specific actions that support herons or through participation in more-encompassing habitat and ecosystem conservation programs. That people and herons have proven over and over again that they can live in harmony is the best hope for the long-term future of this group of birds.
Introduction
Conserving Herons is the action plan for the conservation of the herons of the world. It assesses the conservation status of over 250 populations of herons around the world, suggests a conservation strategy, and summarizes specific conservation projects needed to effect the conservation of the herons around, It is the third version of such a plan (the earlier reports being Hafner et al. 1996 and Hafner and Kushlan 2002). Input is welcome for future versions. It is fundamentally based on two major syntheses of the conservation needs and biology of the herons of the world (Kushlan and Hafner 2000, Kushlan and Hancock 2005) as well as subsequent information from many sources. These two books should be consulted for detailed information and supporting documentation, which are not repeated in the present report.
Conserving Herons is a contribution of the Heron Specialist Group. This Group was founded over 20 years ago by Heinz Hafner and James Kushlan (for its history see Hafner et al. 1986, Hafner and Kushlan 1990, 1996, Kushlan and Hafner 1991, 1993). The Heron Specialist Group is an association of individuals and institutions committed to the conservation of herons. Its sole purpose is heron conservation. It is recognized as the lead worldwide partnership for the conservation of herons by the International Conservation Union (IUCN) and Wetlands International. The Group serves principally as a communication linkage among its over 200 members.
Conserving Herons concerns heron species and population – 62 presently-recognized extant species of the avian family Ardeidae and 259 populations that have been delineated for conservation purposes. Although fundamentally a tropical taxon, herons are well distributed over much of the globe, from the tropics to the far reaches of the temperate zones and even into the sub arctic. Many species are large, visible, human-habitat oriented and, therefore, well known. Others are small, cryptic and retiring and, therefore, little appreciated. Some have large, nearly cosmopolitan, ranges. Others are now restricted, as far as is known, to a few struggling and increasingly isolated individuals. Heron populations often occur within human dominated landscapes. Some have come to depend on human activity for their well-being. Herons have shown themselves to be adaptable. Most populations are not in immediate threat. Some are, however, and these require immediate conservation action.
Not since the 1800’s, when heron populations were decimated in many places in the world because of the plume trade, have herons faced such critical pressures as they do today. Some species and populations are balanced on the verge of extinction; a few may no longer be salvageable. Among these are the nine species and 31 populations currently assessed as being at risk. Four populations have already gone extinct in historic times. It is hoped that Conserving Herons will encourage conservation action for the most threatened of the herons, habitats, and important places. Action on behalf of less threatened species is also needed, to keep common herons common, as are global and continental initiatives to organize conservation, manage data bases, and maintain communication linkages. Conservation action on behalf of habitats herons use, especially natural and artificial wetlands, is needed almost everywhere.
Conservation Strategy
Herons comprise a natural taxonomic group of species found around the world, with the greatest diversity being in the tropics. Herons are for the most part relatively large, readily noticed, and well appreciated by most local cultures and societies.
They are obvious large predators where they occur, and so interact with prey also of interest to humans. While typically birds of wetlands and seacoasts, they also use other habitats. Many herons have been able to accommodate to their landscape being usurped and altered by humans. Some populations have become dependent on these human- altered and human-managed landscapes. Conservation of habitats and sites and linkages among sites are the best single approach to heron conservation. However, basic information on status, trends, dispersion, movements, year-round habitat requirements, important sites, and survival are lacking for many species. However appreciated herons are by people, few active conservation programs likely will be undertaken on their behalf alone. Some such programs are critical, to prevent extinction. But in other cases, heron conservation can and should be included within broader conservation programs. These characteristics of herons and their conservation situation inform development of an overall strategy for organizing conservation action.
The global conservation strategy for herons recognizes the need for information, planning, communication, and actions. Actions involve populations, habitats, sites, flyways, and partnerships and are carried out at global, regional and local scales. It is within the framework presented by these strategic considerations, that global and regional conservation projects are set.
• Information: Much is known about herons and their conservation needs. These have been summarized in two volumes (Kushlan and Hafner 2000, Kushlan and Hancock 2005). However much is left to be discovered, in that for many populations the most basic, yet critical, information on numbers, trends, and dispersion is not available. Perhaps surprisingly, species limits are not even understood for some herons. For most of the over 250 populations of conservation interest, sub-specific studies have not been done or are conflicting in their findings. So herons are a family of birds that requires much additional research of evolutionary relationships using molecular techniques in order to better define populations from a conservation perspective. Although some species are very well known, mostly those from North America and Europe, others, in fact the majority, are much more poorly understood and in some case nearly unstudied. Research is needed to produce new information required for conservation. Even very basic studies of habitat requirements, population status, movements, and demography are valuable to conservation. The Plan encourages the research and monitoring necessary to support heron conservation.
• Planning: The Heron Specialist Group, through its conservation plans, systematizes approaches and priorities to heron conservation action. Priority is given to global actions and local and regional actions for populations under threat. Projects are described that address the principal factors adversely affecting these at risk populations. Within the general overall framework and priority projects, heron conservationists and committed organizations work at their own scales and in their own ways to further the overall goal of heron conservation.
• Communication: Heron specialists and conservation-oriented organizations are scattered around the world, but their goals, interests, tools, and problems have much in common. Communication is the key to encouraging standardization of techniques, focusing activities, and sharing results. The Heron Specialist Group is a network for communication among heron specialists. The conservation plan, reports, scientific papers and books, web site, and periodic meetings are mechanisms for accomplishing communication exchange. Similarly important is communication with other conservationists, nongovernmental organizations, governments, and the public.
• Population: Conservation actions on behalf of heron populations are informed by the assessments of population conservation status, which provide ratings of threat, population estimates, and population threshold values. By far, most of the conservation projects identified in this plan are based on the needs of specific populations.
• Habitat: Conservation actions on behalf of heron habitat focus especially on those habitats used by the most threatened populations of herons. There are a few cases where direct interventions in habitat conservation on behalf of herons are required. However, for both herons at risk and other populations, conservation of habitat in most cases should be part of more-encompassing conservation planning and habitat conservation programs. Herons are part of the ecosystems – natural and man made – that support them. It is these ecosystems that must be conserved.
• Sites: Conservation actions on behalf of sites of importance to herons are carried out in large part, although not entirely, through the programs for identifying Wetlands of International Importance and Important Bird Areas, many of which include important heron sites. Thus heron conservation is done within conservation partnerships with the Ramsar Bureau, BirdLife International, national and regional authorities and local site champions. Heron sites are networked through communication linkages.
• Flyways: Conservation actions within heron flyways are carried on in concert with more encompassing flyway programs. Herons for the most part follow the migratory pathways used by other waterbirds and so flyway conservation that encompasses other waterbirds benefits herons as well.
• Partnerships: Heron conservation is inherently a partnership activity. In only a few cases will it be possible or even desirable to launch focused heron conservation action programs, although in those few cases these are essential. Effective heron conservation is carried out as part of more encompassing programs, such as biodiversity protection, flyway initiatives, wetland and coastal conservation programs, continental and national bird conservation initiatives, protected areas conservation, and local conservation initiatives. Partnerships with governments are critically important, and these need to be encouraged to adopt national, regional and local policies favoring heron conservation.
• Global: Some activities are required at a global scale, such as managing planning, communication, and information exchange. The Heron Specialist Group and its sponsors organize and encourage these global activities.
• Regional: Populations are defined in large part by their local distributions. Regional approaches to conservation can effectively focus on local populations in need by targeting the specific issues facing them as understood by local experts. The Plan is organized around projects that are identified on regional bases. Most of these projects are complex and need to be undertaken by networks and partnerships of individuals, organizations, and governments.
• Local: While the Plan is global and regional in its scope, it is not overlooked that all real conservation action is local. It is the local communities who live with herons, local organizations that undertake specific conservation action, and local sites that need to be protected. Sharing information on the importance and methods of heron conservation with individuals, institutions, local authorities, and local educational institutions is needed. Environmental education programs are central to effective local action, and herons have been shown to be useful in conveying a variety of locally focused conservation messages. The use of herons in local environmental education is encouraged. Activities of local communities of people who live with herons are recognized as central elements of heron conservation.
Conservation Approaches
Population Conservation
The Heron Specialist Group assesses the conservation status of populations of herons, which are delineated for conservation purposes both taxonomically and geographically.
The results of the current (2007) assessment is shown in Table 1. This table provides a fundamental organizing framework for the conservation of heron populations. It reflects the results of collaboration among the Heron Specialist Group, Wetlands International/Ramsar Bureau, and BirdLife International/ IUCN. Taxonomy and English names for species are consistent with Kushlan and Hancock (2005) and Delany and Scott (2006) (although not BirdLife (2004)). The order of listing of populations, population estimates, and 1% conservation threshold values are consistent with Delany and Scott (2006). Conservation status of species at risk (shown in bold font) is consistent with the IUCN RedList assessment (BirdLife International 2004, IUCN 2006). Bold entries in the population status column represent subspecies considered by BirdLife (2004) to be full species. Kushlan and Hafner (2000), BirdLife International (2004), IUCN (2006), Kushlan and Hancock (2005), and Delaney and Scott (2006) should be consulted for background and references to the taxonomic decisions and conservation assessments shown in this table.
Three species and one subspecies of herons in this list are likely extinct. These are all island forms which were adversely affected by the impact of human colonization. Nine species are endangered: Malagasy Heron, White-bellied Heron, Slaty Egret, Chinese Egret, Malagasy Pond Heron, White-eared Night Heron, Japanese Night Heron, Australasian Bittern, and New Guinea Tiger Heron. All of the most threatened species occur in the Eastern Hemisphere, none in the Western. Two species are from Madagascar. The night herons and tiger herons are species of tropical forests. Given the level of risk, specific conservation actions plans and programs are required for each of these species. Additional information on the biology and conservation of each heron species may be found in Kushlan and Hafner (2000), Kushlan and Hancock (2005) and BirdLife (2004).
Some individual populations are of conservation concern. Of the 259 populations identified to be tracked for conservation, 31 are considered to be near threatened, vulnerable or endangered – about 12%. Few data on population size for any heron are exact, but many are sufficient to suggest an order of magnitude of the population. In some cases it is clear that populations are decreasing, even if quantitative data are unavailable. Overall, most populations at risk are island populations, and some are now exceptionally small. The 1% threshold is given for populations for which sufficient information on overall population size is available. These are the population levels that may qualify a site important to herons to become a Ramsar Wetlands of International Importance or a BirdLife Important Bird Area.
Habitat Conservation
Overall, habitat protection is the most serious concern for the conservation of heron populations.
Herons use a variety of habitats, including many human-altered landscapes. Although some species are relatively terrestrial, using grasslands and forests, herons more typically use aquatic habitats, where they forage by wading in shallow water. Most species depend on shallow wetlands and on coastal environments – habitats that are under threat everywhere. Because herons are dependent on wetland function, their presence and population status can serve as biological indicators of habitat condition. Many heron populations are migratory, depending during the course of an annual cycle upon a succession of habitats and sites sometimes at great distances apart, even on different continents. The importance of protecting habitat is reflected in the goal to the identify and protect important sites for herons and of connecting these through flyway and communication networks. Conservation of wetlands and other heron habitats, including Important Bird Areas, depends in large part on local and regional social and economic factors. It is local communities, organizations, and governments that must be committed to the sustainable use of their environment. So local action and local initiative are essential to conserve habitat of the world’s herons. As an adaptable family of birds, herons can live in harmony with people provided the habitat used by both herons and people is properly maintained. Characteristic habitats used by herons of conservation concern are shown in Table 2.
Habitats where herons occur are often at risk. Nesting sites particularly need to be inventoried and protected. Feeding and roosting sites used while nesting, on migration, and during non- nesting seasons also need to be identified and protected. There are occasions where populations are at particular risk or habitat is very limited that conservation action is required directly in support of heron conservation. Without action in support of these heron populations, it is likely these populations will not persist. For most species, it is particularly effective for conservation of habitat to be undertaken within a larger land management context, such as wetland protection, watershed protection, flood control and water supply management, green space preservation, biodiversity conservation, or nature reserves. Humans nearly always play a dominating role in habitat used by herons. Much of human activity in habitats they co- occupy with herons is not detrimental to herons, some human activity greatly enhances habitat value, and some activity has become so supportive that heron populations now depend on its continuation. With proper conservation action, much of human activity and heron habitat use can be compatible.
Habitats that herons use are variable and seasonal locally. Temperature or rainfall, or both, vary in ways the effect herons profoundly. Many populations migrate, both temperate herons in response to annual seasonality and tropical herons in response to wet/dry changes. Most temperate populations migrate to warmer climates for winter. Some populations are highly irregular in their movements. In Australia, inter-annual differences between flood and drought years substantially change the extent and dispersion of available habitat. In South America, ocean oscillations change the rainfall and runoff patterns from one year to the next. Any climate change, such as global warming, that effects rainfall can cause significant changes in wetlands and therefore of the patterns of heron use. Conservation of habitat for herons needs to be cognizant of seasonal, annual, and long term changes in weather and climate. The most threatened of the habitats on which herons depend are wetlands, tropical forests, tropical coastal lagoons, islands, reedbeds, and rice fields.
Overall, conserving wetlands is an important element of conserving heron populations of herons worldwide. Since the beginning of the 20th century, wetlands have been drained, canalized, flooded, and otherwise altered. The process of wetland modification is only accelerating. Wetlands that survive largely already have been degraded to some extent. It is of special importance to conserve those few remaining large wetlands, on which multiple species of herons and other waterbirds depend. These include the Everglades and Mississippi valley of North America, the Amazon, Llanos and Pantanal of South America, the Danube and Rhone deltas of Europe, the Sudd, Congo, and Okavango of Africa, the Tigris-Euphrates, Indus, Ganges, and Mekong of Asia, the Fraser-Darling of Australia. The Fraser-Darling and Okavango, for example, support the majority of certain populations at high risk.
In wetlands, schemes for the management of waters and lands have tended to adversely affect herons at every turn. Mangrove forests appear to be under particular threat throughout the tropics. Conversion to agriculture and aquaculture is a particularly serious threat. In Europe, many of the valuable wetlands in East and Central Europe are still unprotected, particularly the great river floodplains, where major threats are still posed by development programs. In Australia’s Fraser-Darling Basin, a long history of man made changes to the natural flows have been detrimental to heron populations, which are further complicated by the extensive inter-annual variation in rainfall. In North America, the largest tropical wetlands, the Everglades has been reduced to half its size and its natural flow regime altered resulting in well documented population decreases in herons. In Mexico and Central America, wetlands have been being drained for millennia. On the smaller islands of the Caribbean, few wetlands now remain, many of these are privately owned, and nearly all are under immediate threat. In the Greater Antilles and SubSahara Africa, large wetlands are essential habitats for both resident herons and those from continents to the north. There seem to be compelling correlations between conditions on African wintering grounds and the status of European breeding populations, suggesting the importance of habitat outside the breeding zones.
Wetland patches are localized. Most wetlands are to some extent ephemeral with size, area, and value to herons varying among years. A site, which contains a multitude of small wetlands in dry years, may become a single large wetland in wet years. Some site are not suitable except in exceptional years. Heron populations using extensive wetlands require an entire localized network of protected sites, some of which may not be used every year. Maintaining the local dynamic nature of wetlands is a key to their functioning and to their use by herons. Where natural hydrological dynamics have been irreversibly altered, active, or at least heron-sensitive, management of wetlands especially water levels and hydroperiods, may be required for heron conservation.
Next to wetlands, lowland tropical forests appear to be the heron habitats most critically at risk. Given herons’ overall dependence on wetlands, it might be unexpected that lowland tropical forests are habitats of such critical interest. But several of the most endangered species are characteristic of this habitat, which is under relentless pressure from logging everywhere. The White-eared Night Heron, Japanese Night Heron, Malayan Night Heron – all in Asia, New Guinea Tiger Heron – in New Guinea, and the White-crested Tiger Heron – in Africa, are species that depend on this habitat. Reduction in extent, connectivity and functioning of the lowland tropical forests is the principal cause for the vulnerable conservation status of these species.
Tropical coastal lagoons and shores are the essential habitats for a several species and ecologically distinctive populations of herons. These sites are characterized by their shallow, warm, clear water, and as such are also attractive to humans who use and abuse the environment. It is in the coastal tropical wetlands and lagoons that some of the greatest threats to herons occur. Development, drainage, conversion, aquaculture, over-harvest of forage species, pollution, and other degradations threaten some of the most important coastal heron sites. Some populations are specifically adapted to these habitats, such as the Great Blue Heron (Great White Heron) and Reddish Egret in the Americas, several populations of the Little Egret, the Chinese Heron, and the Eastern Reef Heron. Because of the intensity of human economic interests, protection and restoration of these habitats are complex both biologically and economically. Due to their complexity, it seems that parts of such zones should be set aside as protected areas, which can support herons.
Islands are critical to herons. Most herons are colonial and nest on islands, either true off shore islands, islands in lakes, rivers, wetlands and floodplains, or distinctive habitat islands within a larger habitat. Nesting and roosting islands provide protection from predators and disturbance and permit aggregative behaviors to occur unimpeded. Human impacts on islands are often profound. Small, isolated islands suitable for nesting are often at risk to development, disturbance, predation, and degradation. Herons themselves tend to degrade their nesting sites. Where islands are in short supply individual sites may require active management. In fact herons accommodate well to the provision of artificial nesting sites on islands protected from disturbance and predation. Large islands and island chains often support ecologically or taxonomically distinctive populations. Several island species/populations have gone extinct in historic times or are at risk today, showing how vulnerable island populations can be.
Reed beds are the essential habitat for herons such as bitterns and the Purple Heron. All the heron populations in Europe that are of concern depend on reed beds for breeding and feeding. Reedbed alteration, due to drainage for agriculture or industry, disturbance by tourism, reed harvest, and pollution, has negative effects on herons. Nature reserves to protect these beds are highly desirable. Active heron-sensitive management of harvested fields is needed. Fortunately much has been learned in recent years to lend guidance to such management.
Flooded ricefields have become critical habitats for herons nearly wherever they occur. In Europe, with the loss of natural habitats, herons have shifted to ricefields and now depend on the continuation of beneficial ricefield cultivation. The favorable habitat provided by ricefields undoubtedly contributed to heron population increases in northern Italy, where much of the western European population of the Black-crowned Night Heron is now concentrated. In Spain, Purple Herons, Little Egrets, night herons, Squacco Herons and Little Bitterns are concentrated in areas of intensive rice cultivation. A similar situation exists in the Axios Delta in Greece, southern Turkey (Goksu and Cukurova Deltas), Nile Delta of Egypt, and the Camargue of southern France. Because herons depend on rice cultivation in flooded fields, modification in the management of these fields, especially conversion to drier cultivation styles, would have serious detrimental effects on heron populations. Rice cultivation also can be detrimental to herons, especially when it replaces natural habitat that is itself highly productive for herons. Also pesticides used in rice cultivation can adversely effect herons. Encouraging and enabling the management of ricefields, worldwide, to continue to support waterbird conservation is a high global priority.
Site Conservation
The dependence of herons on specific habitats, which often are quite localized, provides an important opportunity for conservation.
Important nesting areas can be determined for each population and efforts undertaken to protect and manage these sites. The importance of conserving specific sites for herons cannot be over-emphasized. Most herons breed colonially and so depend on a few specific sites for nesting, as opposed to being widely dispersed across the landscape. Species that do not nest colonially also are site dependent in that they nest in space-restricted habitats, such as wetland patches. Feeding sites are also localized in most areas. Herons either feed where they nest or fly to feed in changeable places within flying distance from the nesting site. Further, herons depend on restricted areas to feed and roost, such as wetlands, ponds, lake edges, small islands, and seashores. Herons also depend on specific areas during migration, and wintering. Heron conservation is particularly benefited by the identification, protection, and management of specific locations.
Two international programs are designed to identify sites that are of critical value to a significant percentage of a bird population. Wetlands of International Importance are identified by nations that are signatories to the Ramsar Convention. Important Bird Areas are identified by BirdLife International and its national partners.
Congregatory waterbirds enjoy special criteria for identifying such sites. Many site designations have been based on numbers of congregatory waterbird species, and these sites usually support herons. Locations designated as Ramsar Sites and Important Bird Areas constitute the fundamental worldwide network of important sites for herons. However, within a country, Ramsar sites are relatively few and these are seldom sufficient for regional heron conservation. Important Bird Areas fill the gap to preserve other sites of importance to herons. Still additional sites that may not qualify as national or global IBAs also may be of critical importance to local populations. The worldwide identification of important areas for herons is central to the global conservation strategy. Identification, evaluation, management, protection and networking of important sites for herons need to be an ongoing process. The sites currently within these networks change regularly, and the most current lists are available on line (www.ramsar.org/index_list.htm, www.wetlands.org/rsis/; www.birdlife.org/datazone/sites/index.html).
Congregating in one place puts multiple individuals at risk. Colonial nesting and communal roosting expose an entire local population to whatever happens at a the site. Humans taking of eggs and young, which still occurs in many areas, is an extreme form of disruption. Historically, colonies were abandoned when adults were shot for their plumes. Predation by feral animals and various forms of disturbance by people similarly cause nesting failure. The situation is aggravated by rapid industrialization, urbanization, and tourist development, which brings people in closer contact with congregatory sites. This is especially critical in areas of economic deprivation and human population expansion. Because of this sensitivity, colonies, roosts and concentrated feeding sites need to be protected from disturbance and alteration. The efficacy of such protection depends in large part upon the local customs and habits. Cultures that cherish wildlife have protected colony sites for centuries. Cultures that eat wildlife tend to place colonies at grave risk, as heron nestlings are seen as quite edible. Cultures that shoot birds for sport, often include herons as targets. Cultures that emphasize development destroy important sites. Most species of herons can accommodate to repeated, non-damaging disturbance, as can be seen in many places, where herons nest in urban parks, farms and suburban settings. Many species are tolerant of high levels of human activity so long as there remain suitable feeding sites and any disturbance is persistent and non intrusive.
Congregating in one place also provides the opportunity to focus conservation action at a few sites. This allows conservation to emphasize identification, inventory, protection, and management of sites over long periods within a regional landscape. Congregation sites also provide excellent tools for local conservation education.
Flyway Conservation
Most temperate zone species of herons are migratory, as are many tropical herons, which also undertake seasonal movements.
Thus for their year-round conservation, most populations of herons require a seasonal succession of habitats. Four of the five most vulnerable heron species in Europe (Little Bittern, Black-Crowned Night Heron, Squacco Heron and Purple Heron) are trans-Saharan migrants. Drought in their wintering area is probably a factor in the widespread decline of the European breeding populations since the 1970’s. These populations can be conserved only by protecting the web of nesting sites, feeding sites, roosting sites, stop over sites, and wintering sites used during the year. Flyway conservation is a mechanism to bring together heron conservation activities across nations and even continents. The Convention on the Conservation of Migratory Species of Wild Animals (Bonn Convention) provides one mechanism to encourage the conservation of herons and other waterbirds on a flyways basis. This convention aims for the conservation and effective management of wild terrestrial, marine and avian species, especially those considered as “endangered” over the whole of their migratory range. In Europe, under the Bonn Convention, an Agreement on the Conservation of African-Eurasian Migratory Waterbirds (AEWA) covers an area of about 60 million square kilometers, encompassing more than 116 range states in Africa, Europe and Asia. This agreement incorporates eight heron species. Other flyway agreements and initiatives are encouraged as a way of protecting heron migration routes. Flyway approaches to the conservation of herons and other waterbirds, outside the convention can also serve to organize widespread actions. For heron conservation globally, Ramsar sites and Important Bird Areas of importance to herons may be best organized on a flyway basis.
Global and Regional Conservation
Heron conservation has much in common world wide. Global and regional conservation plans, programs, and actions provide support, guidance, continuity, and consistency. Maintenance and enhancement of the Heron Specialist Group network provide networking among specialists. Periodic gatherings of specialists are needed to summarize status, conservation, and biology as information expands.
To determine global population sizes, cooperation is needed among individuals and organizations involved in inventory and monitoring range wide. A global database would allow evaluation of status and long term trends. Collaboration with site networks, conservation assessments, and population estimations need to continue. Regionally, nationally, and locally, specific projects are needed to provide conservation action for species of special concern. Below is a listing of priority conservation projects, with a brief description of the project goals, project rationale, and a brief project statement.
Global 1. Global strategy for inventory and monitoring of heron nesting populations
Goal: Consistent inventory and monitoring activities of heron populations at selected important sites with data archived in a readily available web-based data base.
Population numbers and trends of herons are very little understood for most species and populations of conservation concern. Yet accurate population and trend estimates are essential in order to target conservation action where it might be most effective. Determining population levels and trends, assessing status, and identifying Important Areas need to be ongoing processes. Data should be archived in ways that make them readily available. Inventories of nesting populations and sites should to be undertaken in each appropriate political entity by local stakeholders. Depending on resources available, inventories should at least attempt to achieve a population estimate accurate to an order of magnitude. Ideally population inventories should be accurate to within 20% of actual. Based on inventory information, monitoring programs should be developed on a regional basis (political or ecological) and strive to have a sufficient accuracy to determine population size within 20% of actual and with sufficient precision to be able to detect a rate of population change equivalent to a 50% decrease over a period of 10 years or three generations. Standard methods of census and analysis should be used. Monitoring programs require a determination of bias, which then permits calculating actual population size and trends using data from multiple surveys. Few existing waterbird monitoring programs meet these standards, and in many cases entirely new programs should be developed. Data must be reported and archived in an international database. Selection of such a worldwide data base should be a high priority.
Organize a global program of inventory and monitoring of heron nesting populations and important sites, according to standard protocols, and archived in a universally accessible way.
Global 2. Global network of important sites for herons
Goal: A network of critically important sites for herons worldwide.
The Ramsar Bureau and BirdLife International are engaged in ongoing identification of sites of importance to congregatory waterbird species. Herons benefit from many such sites, even if their populations are not used to satisfy threshold criteria. However, herons also can also serve as reasons to identify such sites. Heron biologists and conservationist should work toward nominating sites of value to herons for listing as important bird areas and Ramsar sites. Sites already listed should be evaluated for their importance to herons and those so deemed should become part of the international heron site network. Information on important heron sites should be made available via internet. Management tools should be made available for sites to use.
Identify a global network of important heron sites; provide conservation tools.
Global 3. Long-term monitoring and research on herons at important sites
Goal: A network that monitors and investigates herons in important sites using compatible methodologies.
The network of sites important to herons (IBA/Ramsar) provides a framework for in depth study and monitoring of herons. Site-specific information on changing population numbers, habitat use, and ecology of herons is required in order to propose appropriate management for important sites. Comparative information on relationships among heron population size, conservation status, foraging and nesting habitat quality, and environmental change can lead to generalities that are transportable between sites. Many ecological relationships can only be determined through long-term studies. The global network of heron studies especially should include the most important heron sites, especially those in large wetland complexes such as the Everglades, Okavango, Amazon, Orinoco, Pantanal, Tigris-Euphrates, and Sudd.
Develop consistent monitoring and habitat use research programs at important heron sites.
Global 4. Tools for heron conservation
Goal: A handbook of standard procedures useful for heron conservation action.
Herons and their important places are widely scattered over the world. As a result those who have responsibility for such populations and sites are similarly scattered and not easily in communication. It is often difficult to find information on how to positively affect herons. However, such tools do exist, having been perfected over the years at many places. It would be helpful to assure that these methodologies are readily available to heron conservationists, preferably through websites.
Collect and synthesize successful methodologies useful for heron conservation and make these available.
Global 5. Taxonomic studies of the herons of the world
Goal: Better understanding of the evolutionary and systematic relationships of herons.
Understanding the evolutionary relationship among heron populations is required to develop conservation programs that place appropriate emphasis on distinctive populations. Recent molecular studies have shown unexpected results at species and higher taxonomic levels. Such molecular studies may be the only way to unravel the relationship among disjunct, similar looking herons. For example, as recently as 2005, Kushlan and Hancock realized that the large white egrets of Asia were in fact two species. Twenty years before, they proposed that herons previously considered to be as many as four species be considered a single species, but there is as yet no molecular data with which to test this hypothesis. Genetic information is needed on other heron species and populations as well. Some of the studies needed include the Butorides complex, the Ardea alba/ modesta/ intermedia complex, Ardea herodias/ wardi/ occidentalis/ fannini complex, Ardea cinerea/ monicae/ sumatrana complex, Egretta garzetta/ dimorpha/ gularis complex, Cochlearius cochlearius complex, Ixobrychus exilis complex.
Conduct taxonomy studies of selected heron populations using molecular techniques.
Global 6. Partnerships in heron conservation
Goal: Insure heron conservation has a place in comprehensive conservation initiatives.
Worldwide, conservation of herons can take place most efficiently within the context of more comprehensive initiatives. Flyway initiatives, all-bird conservation initiatives, continental, national and regional conservation planning, landscape and wetland conservation programs, and International Conventions such as Biodiversity, Wetlands of International Importance, and Migratory Species are examples of large scale conservation programs from which herons benefit. Locally, regionally, nationally, and continentally, heron conservationists should partner and engage these larger conservation programs to meet the conservation needs of herons. Specifically herons should be featured in flyway initiatives in Europe and Asia. A waterbird flyway initiative that includes herons is needed in the Americas. Wetland conservation programs are underway in many locations in the world; herons should be a critical component of such programs. In North America, several bird conservation initiatives include herons or their habitat within their scope. The hemispheric initiative, Waterbird Conservation for the Americas, includes herons within its conservation mandate. These trends of including herons in wider conservation efforts should be encouraged and expanded whenever feasible and appropriate.
Imbed conservation of herons within comprehensive conservation programs and initiatives.
Global 7. Captive breeding of heron populations at risk
Goal: Manage captive stocks of herons worldwide to achieve greatest conservation benefits.
Herons are kept in captivity in zoos and other facilities around the world. There is important education value of such displays to the local community. Stocks are, and should continue to be, managed to maintain genetic diversity and long term viability of the captive populations. The Ciconiiform Taxon Group partners with the Heron Specialist Group to assure proper management of captive stocks. It is important to focus husbandry on the most critically endangered species and populations, both to preserve the species and for eventual repatriation when habitat and ecological conditions merit. Herons that are candidates for captive propagation include White-eared Night Heron, Japanese Night Heron, White-bellied Heron, Cape Verde Purple Heron, Asian Goliath Herons, and island populations such as those from Madagascar.
Maintain registry of herons in captivity; focus husbandry on populations at greatest risk.
Global 8. Bibliography of herons
Goal: Long term availability of the bibliography of herons.
A bibliography of literature on herons has been prepared and is available on line. It is up to date as of 2000, but has not been further updated. The bibliography should be updated. It should be re-designed so that readers can input new entries directly into the data base. A secure, long term sponsor needs to be identified.
Update and maintain long term the on-line bibliography of herons.
Global 9. Worldwide distribution of herons
Goal: Correction and evaluation of the distribution of herons worldwide.
A recent publication updated information on the distribution of all heron species of the world. Maps showing these distributions have been available on the Heron Specialist Group web site. Although these maps relied on the best available information, they are clearly not correct in all cases. For some species, the details or distribution are not correct; however for others it is likely that the ranges are seriously incorrect in places. It is important that information on ranges of heron species be as correct as possible. Ranges as they are currently depicted should be further evaluated by local experts and new information collated as it appears. A system should be devised so that changes to published ranges can be proposed on-line. The patterns of heron distribution worldwide should be analyzed for patterns of biological and conservation significance.
Update current range maps and further analyze distribution of the herons of the world.
Asia
Asia is home to a diversity of heron species and populations and to several of the most endangered herons in the world.
Deterioration of critical habitats, particularly coastal swamps and lowland forests, has effected these populations. Little information exists about some of these species, and their population status and important sites are hardly known. More information on distribution, identification of important sites, and conservation planning are essential for these endangered and vulnerable herons. Conservation in all cases requires the engagement and commitment of local communities and authorities. Basic information is needed for most species. Inventories and monitoring programs need to be developed in many areas, some of which are nearly inaccessible. A few species are in such dire need that captive propagation is required. Several species are teetering on the verge of extinction; action on their behalf is essential, now.
Asia 1. Conservation of the White-eared Night Heron
Goal: Save White-eared Night Heron from extinction.
The White-eared Night Heron is one of the most, if not the most, endangered heron species. It is known from 17 sites in China and one in Vietnam. All of these sites have been identified since 2001, when the species was first rediscovered – while being sold for food in a market. The Vietnam observation is the first in that country in 30 years. In China, the Oriental Bird Club, Kadoorie Farm and Botanic Garden, Guangxi University, Da Ming Shan Nature Reserve, and Guangxi Forestry Department are among the partners working to save the species from extinction. The total species population is uncertain and, in fact, its status is a matter of some disagreement among knowledgeable conservationists. Its population is certainly fewer than 1000 birds, and likely fewer than 250. Based on the success of recent surveys in finding new sites, these should be continued and expanded to determine the bird’s presence in other sites within its potential range. Recent research has been able to better define habitat use, a combination of mountains, forest (both broad-leaved and mixed forest) and nearby aquatic habitats such as reservoirs, streams, ricefields or ponds. Clarification of its annual movements is needed. Wherever this species occurs, it must be completely protected from forest destruction, disturbance, hunting, and other population pressures from people. Establishing a captive breeding program should be accomplished as soon as stock becomes available.
Better understand population size of the White-eared Night Heron; identify and protect every site used.
Asia 2. Inventory and conservation action plan for the White-bellied Heron
Goal: Finding new sites where the White-bellied Heron occurs; protection of every occupied site.
The status of the White-belled Heron is very unclear, but it certainly
is one of the most endangered of herons. It is known only from two sites in Bhutan and one in India. The population is fewer than 1000, and likely fewer than 250. It has declined over almost all its range and surveys have not been able to locate it in other places. The known birds are so few as to be critically endangered. Additional surveys are needed to determine, especially in Myanmar, if there are other sites in which the species occurs. The Royal Society for the Protection of Nature and BirdLife International have been leading the efforts to study and conserve the known birds and inventory new sites. The species appears to use river forests and avoids human. Every nesting site is critical, and protection must be provided to every site identified. Its migratory movements and year-round habitat needs are nearly unknown, and need to be studied.
Determine nesting locations and seasonal movements of the White- bellied Heron; prepare a conservation plan for the species; protect every nesting site.
Asia 3. Distribution and status of the Japanese Night Heron
Goal: Save the Japanese Night Heron from extinction.
Japanese Night Heron is rare and highly endangered. Its population size is extremely low, and likely declining. There have been few recent confirmed breeding records (only at six sites). The only records of this species outside the breeding season are few and isolated. Clearly what is needed is clarification of the current distribution during and outside the breeding season to identify sites critical to its survival. A better population estimate is needed to serve as a basis for determining conservation status and to help develop the needed monitoring program. A quantitative inventory throughout the breeding range is critically needed. There is also a need to better characterize the sites and habitats used by the species for nesting and foraging. Habitat protection is most important. The Wild Bird Society of Japan has provided a basis for establishing a network of forest reserves for this species in Japan. The confirmed and possible breeding sites should be surveyed to re-confirm the night heron’s presence, and the breeding sites and associated feeding sites that
do exist all need to be protected. The species should be accorded full legal protection in Japan. Wider inventories of wintering areas also are needed. Migration occurs through the Ryukyu Islands and there appears to be a regular wintering population in Taiwan. Protection of remaining forested areas in these two areas would contribute significantly to the viability of the existing populations of this species. A conservation plan should be prepared that focuses on its important breeding, feeding, and wintering sites. If stock is secured, a captive breeding population should be established to eventually enhance the population through future releases.
Clarify the current status, distribution and numbers of this species during and outside the breeding season; protect all sites at which it occurs.
Asia 4. Conservation of the Chinese Egret
Goal: Better understanding of the distribution and protection of important sites for the Chinese Egret.
The Chinese Egret is a vulnerable heron, which has been a species of concern for many years. Due to this concern, much progress has been made in documenting additional nesting sites, stop over sites, and wintering areas, all of which need to be protected throughout its range. Its population is thought to be about 2600-3400 birds, but likely is larger. A coordinated range-wide inventory of nesting colonies and critical wintering sites is now needed. Emphasis should be in China and also areas to the north of the known range, such as Sikhote-Alin and Kamchatka. Basic studies and reviews of the biology of the species are needed to clarify its specific ecological requirements. A network of important nesting and wintering sites should be developed. A conservation plan compiled by all stakeholders should be developed that encompasses every important site. A range-wide inventory of wintering areas is also needed, including those known now to be important (and qualifying as IBAs): Kanghwa Island and Yongjong Island, South Korea, Jiangsu and Yancheng Coasts, Dongting Hu NR, Pangzhai and Quingxu/Huayan Res. P.R.o.China, intertidal mudflats on Palawan and at Ormoc and on Orlango Island in the Philippines, Tapei estuary, and coastal mudflats in Taiwan. Recent studies have found important sites in Malaysia as well. Governments need to be encouraged to protect the known breeding, stopover and wintering sites. A range-wide monitoring program is needed.
Inventory breeding range to find nesting sites; develop a conservation plan that networks these sites and provides protection for the heron.
Asia 5. Distribution and status of Sumatran Heron
Goal: Clarification of the current status and distribution of the Sumatran Heron.
This species appears to be declining throughout much of its range and to have disappeared from most of the mainland south-east Asia (except parts of Indochina) in the last fifty years. A robust population persists in Indonesia, with small numbers in mainland south-east Asia and the Andaman Islands. A primary need is to clarify population size and status of the Sumatran Heron throughout its range in Asia. It is essential to determine the overall numbers of the species and also to refine identification of those populations of conservation concern. Its large size and solitary nature makes surveys and monitoring difficult, so a network of conservationists will be needed to determine its status and distribution and attend to its long-term conservation.
Clarify the current status and distribution of the Sumatran Heron in Asia; identify and assess the viability of its populations.
Asia 6. Inventory and status of Grey Heron population in Sumatra
Goal: Understand long-term viability of the Grey Heron population in Sumatra.
This population of the Grey Heron is restricted to the island of Sumatra. Very little information is available on waterbird populations on the island, and so little is known about this heron. The population is thought to be about 1000-2000 birds and is considered to be vulnerable, It occurs over a restricted range of habitats that are under threat. An inventory of the population needs to be undertaken to determine population size and nesting sites. The appropriate taxonomic status of this population needs to be determined.
Conduct an inventory of the Grey Heron on Sumatra; determine important sites; examine its systematics.
Asia 7. Status of Schrenck’s Bittern
Goal: Understand status of Schrenck’s Bittern during and outside the breeding season
The status of this species is not well known. It is necessary to clarify the range and location of sites of importance for this species outside the breeding season by conducting a thorough survey throughout its known distribution. Breeding season surveys should establish a more precise estimate of breeding populations. Similarly its non-breeding range needs to be better understood. Although the main non- breeding distribution likely is in the Philippines, there are records from various sites and habitats throughout Southeast Asia. Surveys should be undertaken of sites where the species is known to occur (concentrating on the Philippines) to identify the main non-breeding distribution. Information collected should be gathered and a conservation plan devised including, as appropriate, site protection of important nesting and non-nesting areas and hunting control.
Conduct an inventory of breeding and wintering areas of the bittern; develop a conservation plan.
Asia 8. Distribution of Indian Pond Heron in the Bay of Bengal
Goal: Understand status of the Bay of Bengal population of the Indian Pond Heron.
The waterbirds of the islands off the coast of Myanmar in the Bay of Bengal are little known. The Indian Pond Heron is one of these. It is desirable to develop and implement an inventory of the Indian Pond Heron in the area. This information would provide the basis for implementing waterbird conservation in the area.
Conduct inventory of the status of the Indian Pond Heron in the Bay of Bengal near Myanmar.
Asia 9. Conservation status of the Indian Pond Heron in the Maldives
Goal: Better understand status of the Maldives population of the Indian Pond Heron.
The Maldives is a large and complex system of islands that can be very difficult to access. Little is understood about the status of the Indian Pond Heron population of the islands. It is considered to be vulnerable. In the short-term, the most important action should be to determine the current size of the population and conduct an assessment of habitat availability and the degree of threat to breeding and foraging habitats. Public awareness may have the greatest potential to improve the conservation status and the habitat, so increasing public awareness may become the main thrust of the conservation strategy.
Conduct inventory to determine population size and important sites for the Indian Pond Heron in the Maldives; increase public awareness.
Asia 10. Conservation status of the Black Bittern
Goal: Better understand the status and distribution of the Black Bittern.
The Black Bittern is extremely poorly known. It appears to be a species of lowland riverine forests. Details of distribution and breeding status are little understood. An inventory should be undertaken to provide information to form the basis for monitoring, carrying out of ecological studies, and developing as a conservation plan for this species.
Inventory the nesting range of the Black Bittern to determine status and conservation needs.
Asia 11. Conservation status of Malayan Night Heron populations
Goal: Better understand the conservation status of populations of the Malayan Night Heron.
Despite its wide distribution and apparent abundance in certain locales, due to its nocturnal habits and use of dense humid forests, the Malayan night heron remains little known. The best information on the species is from excellent studies of the breeding population in Taiwan. Overall, it appears to be substantially less at risk than the other Gorsachius night herons. But given conservation issues with the other species, clarification of the status of this species in various parts of its range is needed. Particularly, a range-wide inventory of nesting is needed on Sumatra and Peninsular Malaysia, where recent records suggest that these areas may support resident populations, Nicobar and Palawan where populations apparently are sedentary (birds on Palawan was sometimes considered a subspecies and are considered to be vulnerable), and also the apparently healthy populations recently located in Cambodia, Laos, Thailand and Vietnam. There is a need to increase survey efforts on remaining suitable forests in Philippines, its wintering range, which is under threat from logging. As there are several apparently disjunct populations of this species, some of which have been considered subspecies, so a systematic study is needed to define conservation populations.
Conduct a range-wide inventory of the status of the Malayan Night Heron in both potential breeding and wintering sites; clarify distribution and the status of its various populations; conduct taxonomic studies of populations using molecular techniques.
Asia 12. Status and distribution of the Goliath Heron in South and Southwest Asia
Goal: Search for populations of the Goliath Heron in Asia.
The south Asian and southwest Asian populations of the Goliath Heron are small, restricted, coastal, little understood, and seldom reported. Nesting status is unclear. If these populations still persist, they are likely to number under 50 birds. As a result the Asian population is considered to be endangered. It is necessary to conduct surveys and develop communication networks to determine the persistence of the species in the region.
Assess the current status and distribution of the two Asian populations of the Goliath Heron.
Asia 13. Distribution of large white egrets in Asia
Goal: Clarification of the distribution of the large white egrets of Asia.
The specific distinctiveness of Ardea alba and Ardea modesta has recently been proposed. Observations by authorities familiar with birds in the field are tending to agree that the two forms are distinctive. Both occur, at least seasonally, in Japan and Korea and have been given different local names, suggesting they are readily recognizable. However distinguishing the field characteristics of the two forms requires additional documentation. It is apparent that the published range maps for the two species in Asia are not correct, as the ranges appear to overlap in winter and perhaps at other times as well. The range of the Intermediate Egret is also unclear over this part of its range. What is needed is a determination of the seasonal ranges of the two large egrets and the Intermediate Egret in east Asia.
Determine seasonal distribution of the Great Egret, Eastern Great Egret, and Intermediate Egret in east Asia.
Asia 14. Conservation of the Eastern Reef Heron in China
Goal: Protection of important sites for the Eastern Reef Heron in Coastal China.
The Eastern Reef Heron, although common over much of its range, is rare and its distribution rapidly decreasing in China. Only a few individuals have been seen on surveys. This species has specialized habitat requirements on rocky or sandy coasts and so comes into conflict with development pressures in these areas. It is essential to determine important sites where the species still occurs and to develop programs to protect these sites.
Survey coastal China to determine sites where the Eastern Reef Heron occurs and develop protection programs.
Asia 15. Inventory of heron colonies in China
Goal: Complete an inventory of heron colonies throughout China.
Recent surveys suggest that China likely supports far more herons and heron colony sites than was appreciated. For example, a new colony of Chinese Egrets with 100 nests was recently discovered in Shandong, and a recent survey of Hainan Province found over
5000 nests of four species. Even larger colonies have been located. Population sizes, distributions, and important sites for herons in China need to be determined. This should be done in multiple ways, depending on available resources, interests and commitments from organizations, such as the Egret Research Group of the Hong Kong Bird Watching Society, universities, and government agencies.
Conduct a thorough inventory of heron colonies in China.
Europe
The European region, including Europe, the Mediterranean, North Africa, and the Middle East, is one of the best studied regions with respect to herons.
Reliable data on heron populations size exist for most European countries and for the most important breeding areas of North Africa. Major gaps in knowledge exist in Syria, Lebanon and Jordan, and in the east Mediterranean. No European heron species is globally endangered, but some populations are at risk, underlying the need for accurate status assessment and protection of important sites.
Europe 1. Status and monitoring of the Eurasian population of the Great Egret
Goal: Better understanding of the status of the Great Egret in Europe, Middle East, and North Africa.
The Great Egret population in Europe is vulnerable due to its small numbers and localized occurrence. However, the population appears to be increasing and expanding in Europe in both breeding and in winter. In Europe, there are few censuses of the breeding population, and only 7000-17000 individuals have been counted in wintering areas. It occurs but does not breed in North Africa. It has been expanding its range in small numbers in a westerly direction. It appears to be most secure in the Eastern Mediterranean, Black Sea regions, and Russia. This Great Egret population deserves expanded monitoring efforts, particularly in the Eastern Mediterranean, the Middle East, Northern Africa, and Black Sea. Monitoring wintering populations is also important, as increases in wintering numbers appear to have presaged increased nesting numbers. Given how localized the breeding sites are, most colony sites require protection.
Inventory Great Egret populations in areas where information is slim; protect all nesting sites.
Europe 2. Monitoring and habitat protection for Eurasian Bittern in Europe
Goal: Documentation of the status and trend of the Eurasian Bittern in Europe and protection of important sites.
The Eurasian Bittern has experienced a notable decrease in west Europe over the past decades, although it remains common in parts of east Europe. In the east, inventories are needed to determine the most important breeding sites. In the west, high densities of this bird are restricted to rare extensive reedbeds, especially in the Rhone delta of southern France. Sites where the bittern occurs in numbers need to be fully protected. The species is rare in North Africa, where breeding only occurs in a few small pockets of favorable habitat. All sites used by the bittern in North Africa need to be protected. New techniques permit more exact estimation and monitoring of breeding populations.
Continue and expand monitoring programs for Eurasian Bitterns in the European region; identify and protect important sites and all sites in north Africa and southern Europe.
Europe 3. Purple Heron conservation and monitoring in Europe, Middle East and North Africa
Goal: Continued and expanded survey and monitoring of Purple Heron in the Mediterranean.
European breeding populations of Purple Herons appear dependent on large reedbeds (>20-30 ha) for nesting, and they forage exclusively in freshwater wetlands throughout the year. They are sensitive to human disturbance. Purple Herons have declined in all European countries except for the small populations in Slovakia, Greece and Portugal. Recently there has been an increase in breeding areas in Italy, France and Spain. Long runs of data exist on the size of breeding populations in the Netherlands, Northern Italy, the Mediterranean coast of France, the Ebro Delta and the Albufera de Valencia in Spain. Purple Herons are rare in the Near East and in North Africa. Purple Heron nesting numbers should continue to be monitored and monitoring expanded to other important sites throughout Europe. To clarify the range in southern Europe and location of the most important wintering sites, a network of collaborators needs to be developed in the African wintering areas. All important sites identified should be protected. Continued collaboration among biologists and conservationists concerned with this species should continue and be formalized where possible.
Continue and expand collaborative inventories and monitoring of Purple Herons; evaluate trends of nesting populations; protect important sites.
Europe 4. Population status and conservation of the Squacco Heron in Western Europe
Goal: Better inventory and conservation action for populations of the Squacco Heron in Western Europe.
The Squacco Heron is the rarest heron in Europe. Its population appears to fluctuate in the western Mediterranean but seems to be increasing at present in southern Europe. It is necessary to continue to monitor breeding populations in the major breeding areas of Western Europe (Italy, southern France and Spain) and to conserve the important sites and wet woodlands. All important sites, as well as many potential sites, in the region need protection. The largest Squacco Heron populations of Western Europe today use ricefields, which are susceptible to changes in agricultural practice. Because Squacco Herons nesting in the Palearctic are long-distance migrants, most of them wintering south of the Sahara, the cause of population decline in Eastern Europe and fluctuations in the western Mediterranean may be operating outside the breeding season. It is necessary to synthesize existing data on habitat use and feeding ecology across the major rice production areas in the Mediterranean region and to prepare management strategies that encourage heron-friendly agricultural practices in rice fields.
Inventory populations of the Squacco Heron in Western Europe; prepare management strategies for protection and maintenance of sites used by the herons, especially rice fields.
Europe 5. Inventory and protection of nesting sites of the Little Bittern in Europe and North Africa
Goal: Securing suitable breeding habitat in as many areas as possible in Europe and North Africa.
The Little Bittern in western Europe and north Africa is considered to be vulnerable because it has strongly decreased both in population size and range since 1970. The species’ range appears to be contracting from west to east. There are few recent breeding records in north Africa. Although it is possible that the decline in European numbers is largely due to high mortality of birds during migration or when wintering in Africa, protection of nesting sites remains essential. Conservation efforts in Europe should concentrate on maintaining favorable breeding habitat over large areas. It is necessary to conduct surveys to determine suitable breeding habitat and breeding status both in the known strongholds of the species and in areas where it has disappeared. Ecological studies need to be undertaken to determine reasons for the decline in breeding numbers in Europe. Surveys should be conducted to determine the most important wintering habitat. It is critical to identify important sites for both breeding and wintering, and protect these sites.
Determine and protect the most important breeding areas for the Little Bittern in Europe and most important wintering sites in Africa.
Europe 6. Trend and habitat requirements of the Black- crowned Night Heron in Europe
Goal: Better protection of breeding populations of Black-crowned Night Heron in Europe.
The Black-crowned Night Heron nesting population in Europe is in slow or moderate decline. The largest, apparently stable, breeding population in Europe is in Italy where it depends on rice fields. Elsewhere, in the southern Mediterranean breeding distribution is limited to a few freshwater areas. Conservation of the species requires continued monitoring of breeding populations in the major breeding areas of western Europe (Italy, southern France and Spain). More importantly, a multinational program is needed for the conservation and management of wet woodlands and reedbeds. Existing data need to be synthesized on habitat use and feeding ecology across the major rice production areas in the Mediterranean region in order to prepare management proposals.
Continue to monitor breeding populations of the Black-crowned Night Heron at important sites in Europe; develop strategies to protect these sites, especially wet-cultivation rice fields.
Africa
For the majority of African heron populations, there are no historical or contemporary quantitative data, and it is therefore not possible to quantify population changes.
Of course, this needs to be rectified. Despite limited quantitative data, it is clear that conservation action for several species and subspecies on the African continent is needed. The Slaty Egret for instance is considered extremely vulnerable due to its dependence on the seasonal wetlands of central Africa. The Eurasian Bittern subspecies in southern Africa is today endangered due to shrinking and deteriorating habitats, and its intolerance of man. The Malagasy region, which includes the island complexes Seychelles, Aldabra, the Comoros, Madagascar, Réunion, Mauritius and Rodrigues, supports two species and eight subspecies that are endemic and require special conservation attention.
Africa 1. Inventory and site conservation for the herons in Madagascar
Goal: Better understanding of the status of all the heron populations of Madagascar and of conservation actions needed.
Among the locations in the world in which herons are in the greatest difficulty, the island of Madagascar stands out. Several species and populations of herons are at immediate risk and they share conservation issues mainly related to habitat destruction and degradation and hunting. The principal habitats are coastal mangroves, riverine floodplains, freshwater swamps and natural lakes. The degree and rate of degradation of aquatic habitats across the island has been alarming. Habitat loss, introduction of non-indigenous fish species, and hunting have caused dramatic population declines many of the region’s waterbirds. Heron species of concern in the region are: Ardea humblotti, Ardea cinerea firasa, Ardea purpurea madagascariensis, Egretta ardesiaca, Ardeola idea, Butorides striatus, Ixobrychus minutus podiceps and Ixobrychus sinensis. A complete nationwide inventory of heron nesting sites is needed to provide accurate population estimates and to identify sites of importance to herons. Sites should be qualified as Important Bird Areas for herons, immediate protection provided, and conservation plans developed for the long term management of these sites. A national monitoring program for heron species at risk is needed.
Inventory the heron populations of Madagascar; identify and protect all important sites; enact conservation actions as needed at each site.
Africa 2. Save Cape Verde Purple Heron from extinction
Goal: Save the Cape Verde Purple Heron.
The subspecies of Purple Heron endemic to the island of Santiago, Cape Verde Islands, is highly endangered. The population has declined significantly since its discovery in 1951, from an estimated 75 pairs to a recent population estimate of 20 or fewer pairs, which use only two sites. Unlike other Purple Heron populations, this subspecies appears to be independent of wetlands, nesting in trees and apparently foraging on arid hillsides. There is no information on foraging habitat outside the breeding season, but it is suspected to use isolated, dry river valleys. The population is not viable under present conditions. Immediate conservation action provides the best possible chance of survival. A complete and accurate inventory of the population and its nesting, roosting, and feeding sites is needed immediately. The government should be encouraged to initiate formal protection of the species from hunting, disturbance and habitat destruction, and to provide immediate protection for the two known roosting/nesting sites. Research should be conducted to obtain and compile information needed to develop a conservation plan for the species, including identifying all nesting sites and foraging habitat and sites used during and outside the breeding season. The main priority must involve survey of all suitable trees on the island (these are very limited in number and therefore this is not as impractical as it sounds) and identification of foraging habitat and sites use. A monitoring program is needed at the two known breeding colonies and any other sites found. Public awareness and governmental commitment to conservation of the subspecies are essential.
Inventory Santiago, Cape Verde, for additional nesting sites; determine feeding habitat use year round; completely protect the birds and every nesting, roosting and feeding site.
Africa 3. Save Eurasian Bittern from extirpation in Southern Africa
Goal: Implementation of conservation action to protect sites used by the Eurasian Bittern in South Africa.
This subspecies of the Eurasian Bittern is confined to southern Africa and is in serious decline due to loss of wetland habitat and its apparent intolerance of human activity. This subspecies is among the most critical heron conservation concerns globally. It is now exceedingly rare or extirpated over much of its former range, and it may be only in Zambia and perhaps the poorly known eastern Angola wetlands that it survives in any numbers. In South Africa it breeds only in Natal and Transvaal, with a total population of fewer than 100 individuals. There appears to be little doubt that the entire population is in serious decline and endangered. The ecology and habitat requirements of this subspecies are little known. It is suspected to require extensive reed or mace beds. Unlike many taxa, the main problem is not a lack of suitable data to show the decline. Consequently, it is vital that urgent conservation action be undertaken. Although there is a need for a better understanding of the status and dynamics of this subspecies, it is far more urgent to implement conservation management and protection of all sites at which it occurs. The main scope of site protection should concentrate on the subpopulations in Zambia, eastern Angola, Natal (South Africa), and Transvaal (South Africa). Censuses should be conducted, using techniques recently developed in Europe. It is critical to address (through liaison with the departments responsible for the environment in the relevant governments) protection of the sites holding this subspecies.
Implement strict protection of each site in which the Eurasian Bittern occurs in southern Africa; clarify the current distribution and population; identify additional sites.
Africa 4. Conservation of the Malagasy Heron
Goal: Protection of the Malagasy Heron.
The Malagasy Heron has an overall population of only 1000-3000 birds and is considered to be endangered due to its localization in a few, threatened sites. It appears hunting continues. It is known to occur and may be still fairly common in four areas in western Madagascar, which provides the opportunity for successful conservation. It possibly breeds also on the Comoro islands, and this needs to be clarified. There is a clear suite of problems, mainly related to habitat destruction and degradation, which affect the survival of a number of waterbird species and endemic subspecies on Madagascar. Hunting and nest predation by locals and conversion of natural wetlands to rice agriculture are persistent threats to the Malagasy Heron. Known nesting sites require immediate and complete protection through efficient wardening combined with public awareness campaigns. It is desirable to conduct an inventory to locate additional nesting sites and potential sites, as well as an inventory to determine status in the Comoros. Conservation will require commitment of the government and conservation organizations active in the Malagasy region to take on the Malagasy Heron as a priority.
Locate and protect all nesting sites of the Malagasy Heron; stop hunting.
Africa 5. Conservation of the Malagasy Pond Heron
Goal: Better define the conservation status of the Malagasy Pond Heron.
Endemic as a nesting species to Madagascar, the Malagasy Pond Heron is considered to be endangered. With a population presently estimated at less than 250-1000 individuals, there has been a sharp decrease over the last 50 years, due to habitat loss and degradation, especially conversion of natural feeding areas for rice-growing where it appears to be out competed by the more recently established Squacco Heron. It is difficult for most observers to distinguish this species from local pond herons in winter. However recent evidence suggests that it may be more common than previous thought in winter in east Africa, suggesting its population is larger than presently estimated. A synthesis of available information and inventories of nesting sites and wintering areas are needed to determine population size. It is critical to identify important nesting sites and protect them. Conservation organizations in Madagascar are urged to take the lead and to collaborate on habitat conservation for the species in Madagascar. Establishing a captive breeding population may be of value. A better understanding of wintering range and habitat use in Africa is important, and this requires a better clarification of field identification.
Identify and protect all nesting sites of the Malagasy Pond Heron; better delineate the wintering range in Africa.
Africa 6. Conservation of the Grey Heron in Madagascar and related islands
Goal: Inventory and protection of nesting sites of Grey Heron in Madagascar, Seychelles, and related islands.
The Grey Heron of Madagascar is a recognized subspecies that is endangered due to persistent threats, principally from habitat destruction, hunting, and predation at breeding colonies. There are thought to be fewer than 5000 birds in the population, and likely far fewer. The taxonomic status of similar herons in the Comoros, Aldabra, Amirantes, and Seychelles is uncertain and needs to be clarified. All nesting sites supporting more than 50 individuals need to be protected. It is desirable to start public awareness programs that include heron conservation in the message.
Identify and protect important nesting sites of the Grey Heron on Madagascar; determine taxonomic and conservation status in Seychelles, Comoros, Aldabra, Amirantes, and Seychelles.
Africa 7. Conservation of the Purple Heron in Madagascar
Goal: Protection of the Purple Heron population in Madagascar.
The Purple Heron race endemic to Madagascar is vulnerable. It depends on freshwater lakes and rivers. Except in the south, it is considered to be common but appears to be declining overall. There is continuing pressure on wetland habitats in Madagascar, where the Purple Heron occurs. The population needs to be monitored and its important sites identified and protected from alteration and disturbance.
Develop monitoring programs for the Purple Heron population in Madagascar; protect important breeding and feeding sites.
Africa 8. Conservation of the Grey Heron in Coastal Mauritania
Goal: Better understanding of the dispersion and taxonomy of the Grey Heron in Mauritania.
The Grey Heron of coastal Mauritania is unusual due to its pale coloration, ground nesting, and extremely restricted range. This small population (7500-12500 individuals) is confined to the Banc d’Arguin in Mauritania and subject to considerable fluctuations in numbers of breeding birds. Due to its restricted range, this population is considered to be vulnerable. The breeding islands are extremely difficult to access and they are part of the Banc d’Arguin National Park, which has strict regulations. Tour du Valat in France and the National Park in Mauritania have collaborated to conduct aerial breeding censuses, which need to be continued. However, the precise distribution and size of this population are unclear. The feeding areas are very extensive and human population is so low that there is practically no disturbance to these birds. The primary aim of more extensive survey work should be to determine the full range, accurate population size, and assess the needs for active conservation. The taxonomic status of this population is far from clear, as there are suggestions that this population may be a relict that is worthy of a higher taxonomic classification.
Determine dispersion and habitat use by the Grey Heron in Mauritania during and outside the nesting season; insure that all important sites are protected; assess the taxonomy of this population using molecular techniques.
Africa 9. Conservation of the Little Bittern in Madagascar
Goal: Better understanding of the distribution and population size of the Little Bittern in Madagascar and implementation of conservation actions.
The Little Bittern population endemic to Madagascar is effected by loss of wetlands on the island. As a result the population is considered vulnerable. As far as is known, it is restricted to a few sites. Identification and protection of these sites is an absolute priority. The population size needs to be better documented and a monitoring program conducted.
Inventory nesting sites to determine important areas and population size of the Lest Bittern in Madagascar; protect these areas; monitor the population.
Africa 10. Systematics of heron populations on the Seychelles
Goal: Better understanding of the taxonomic status of the heron populations on the Seychelles.
Little is known of the taxonomic status of the isolated populations of herons on the Seychelles. These may represent colonists from Madagascar, and may or may not be distinctive. Limited habitat on the Seychelles means that all sites need to be protected. Fortunately a new emphasis on wetland conservation is being shown by the government. It is of importance to understand the genetic distinctiveness of the populations there.
Conduct studies of the systematics of the disjunct heron populations on the Seychelles to determine conservation status.
Africa 11. Conservation of the White-crested Tiger Heron in West Africa
Goal: Better understanding of the distribution and habitat requirements of the White-crested Tiger Heron in West Africa and protection of the tropical forests of West Africa.
Nearly nothing is known about the biology, status or conservation needs of the White-crested Tiger Heron. This species, the only tiger heron in Africa, occurs in tropical forest swamps, is very secretive, has been little reported, and nearly unstudied. It is found sparsely distributed in the west African rainforests of Gabon, lower Congo, and northern Democratic Republic of Congo, although it has declined in other parts of the Congo. It has recently been reported from over 60 sites, suggesting that it may be widespread in suitable habitat. Collecting information on additional sites is essential to help understand its distribution, status, and trend. Basic information on its biology is needed. Clearly protection of the lowland tropical forests of West Africa is essential to the conservation of this species.
Collect information on the distribution and biology of the White-crested Tiger Heron; encourage protection of forests in which it occurs.
Africa 12. Status and conservation of the Slaty Egret
Goal: Protection of important sites and a thorough inventory of the population of the Slaty Egret.
The Slaty Egret is found only in the swamps and floodplains of the major river systems of Central Africa, in Zambia, northern Botswana and Namibia. It is poorly known biologically. Its population is small, and given threats to its restricted wetland habitat, it is considered to be vulnerable. Flood regulation has already caused it to disappear from one part of the Kafue Flats in Zambia, and the population is threatened by a variety of proposed water management actions, such as plans to harness the waters of the Okavango Delta. Plans to clear large areas of the Okavango Delta of tsetse fly may seriously affect the ecology of the entire area and reduce prey for the egrets. Spraying may also disturb nesting colonies. The proposed development of a rice-growing project in the eastern Caprivi, Namibia, may markedly alter the functioning of the Zambezi floodplain. The impact of tourism and livestock industries together with the constant reduction of potential breeding sites by reed-cutting and fires are all likely to have negative effects on its population. Data on populations of this species, while locally limited, appears sufficient for selection of sites for protection, such as the Kafue Flats, Liuwa Plain and Bangweulu Swamp, Zambia; Okavango Delta, northern Botswana; Zambezi Delta, Mozambique; Caprivi strip, northern Namibia. It is critical to establish these as important sites and to implement their protection. A monitoring program is needed to assess impacts and assure conservation measures are adequate. Ecological studies are needed to determine the dynamics of colony-site choice and site fidelity, foraging site selection, habitat-preferences, colony size, and populations. Population status needs to be determined throughout its range in both nesting and non-nesting seasons. Based on the inventory and existing information, important sites need to be recognized.
Identify and protect important sites; increase understanding of the distribution and conservation status of the Slaty Egret.
Africa 13. Status and trends of the Black Heron in Africa
Goal: Better assessment of the existing population size and trends of the Black Heron and better understanding of its biology.
Little is known about the population status and trend of the Black Heron in Africa. It is uncommon and there is suspicion that it is declining on the continent, although quantitative data are few. It is important to develop a survey program to better determine the current conservation situation. Information on its biology is needed, especially understanding relationships between fluctuations in local populations and water levels or rainfall. A monitoring program should be developed.
Assess the existing population size and trend of the Black Heron; better understand its biology.
Africa 14. Conservation of the Black Heron in Madagascar
Goal: Arresting population decline of the Black Heron in Madagascar.
The population of the Black Heron is in serious decline in Madagascar, but its status is not well documented. There is a need to better understand the status of the Black Heron in Madagascar, begin a monitoring program, locate important sites, and begin conservation measures.
Assess the existing population size and trend of the Black Heron in Madagascar; institute conservation measures at important sites.
Africa 15. Status and ecology of the Rufous-bellied Heron
Goal: Better understanding of existing population size, biology, and conservation objectives of the Rufous-bellied Heron.
The Rufous-bellied Heron is local and uncommon. Breeding is apparently sporadic in response to fluctuating water levels. It appears to be declining. However, existing data are not sufficient to confirm this. This species may well be of higher conservation concern than its current conservation status suggests. It is urgent to update a baseline population estimate and distribution from all available information. In addition surveys are needed to improve knowledge of the current population status and identify priority sites for conservation. So little is known about the biology of the species that detailed ecological studies are desirable, especially to achieve a better understanding of its relationship with water conditions.
Synthesize available information; carry out an inventory to assess the current population trends and to identify priority sites.
Australasia
The continent of Australia, the large islands of New Zealand and New Guinea, and thousands of small islands of Oceania scattered across the Pacific Ocean support important heron habitats.
This huge and diverse area is considered as a single heron conservation region. On the Australian continent, the climate fluctuates from extreme drought conditions to extreme flooding. Large wetland systems, the Fraser-Darling and Kakadu, historically supported large numbers of herons. Of the native herons, only the Cattle Egret has been extensively studied in Australia. Data on other heron species are fragmentary. Very little research has been carried out in any of the island countries. Herons are known to move between Tasmania and the mainland of Australia and between Australia and Papua New Guinea. In many areas of this extensive region data are too sparse to truly understand the status and conservation issues of the herons there.
Australasia 1. Conservation of the Great Egret in New Zealand
Goal: Maintaining protection of known nesting colony and better understanding of important feeding areas.
The total population of Great White Egret in New Zealand is about 100 birds and so is considered to be vulnerable. The viability of such a small population is uncertain and critically dependent upon environmental and anthropogenic influences. Although it is encouraging that the population appears healthy and currently to be increasing, this small population must continue to be carefully monitored and its population dynamics and limiting factors studied. The colony and surrounding feeding areas are being guarded, and this needs to continue. It is important to inventory apparently suitable habitat during the breeding season to locate non-breeding birds and to assess the possibility that other colonies exist. Information is needed on feeding areas used during and outside nesting season to identify important sites. Research is also needed into the migration patterns and habitat usage along migratory paths.
Continue to protect the known colony site; survey other potential habitat for additional colony sites and for feeding habitat use year round.
Australasia 2. Conservation of the Australasian Bittern
Goal: Better understanding of the status each population of the Australasian Bittern and identification and protection of important sites.
The current population of the Australian Bittern is not well understood. Overall its populations is estimated to be 2500-10000, the range suggesting the wide confidence limits on the estimate. In Australia, it is thought to number not more 2500 birds. But no site is known to have more than 100 birds, and most recently reported sites hold fewer than 10 birds. Year to year variability in populations size due to differing environmental conditions is not well known, but could be significant to its long term population trend. Clearly more accurate monitoring is needed. Yet despite this uncertainty, it is undoubtedly decreasing in Australia and Tasmania as its habitat is altered. As a result, the Australian populations are considered vulnerable. In New Zealand, New Caladonia and the Loyalty Islands populations are smaller and considered to be endangered. Better data are needed to verify present status and distribution. Important sites for its conservation need to be discovered throughout it range. In each part of its range, awareness should be increased and information provided to local people. Throughout its range strict protection of the species and sites needs to be implemented.
Conduct inventories of the population status and important sites for all populations of the Australasian Bittern; assess trends; protect all important sites; implement additional conservation actions as determined.
Australasia 3. Status of the New Guinea Tiger Heron
Goal: Better understanding of distributions, habitat requirements, important sites, and conservation needs of the New Guinea Tiger Heron.
The New Guinea Tiger Heron is a bird of forested streams, confined to Indonesia and Papua-New Guinea. The species is considered to be near threatened. There is no population information on this species, and no information on its basic biology. In fact only thirty confirmed sightings have been documented in the last fifteen years, and it is currently known to be constantly present at only one site. It is likely more widespread than appreciated but is apparently absent from what should be appropriate habitat. It is among the least known herons in the world. Even the most basic data on its conservation status are needed. This heron is suspected to favor broad, heavily forested lowland rivers, a habitat throughout its range that is under extreme pressure from logging and pollution. The most urgent action needed is to clarify the current details of distribution, provide a population estimate, and carry out ecological studies into its habitat requirements. Protection of the remaining forested lowland rivers is likely a requirement for the species.
Clarify the current distribution of the New Guinea Tiger Heron; provide better estimation of breeding populations; determine actions needed for conservation of lowland river forests.
Australasia 4. Persistence of the Little Bittern in New Zealand
Goal: Clear determination of persistence of the Little Bittern in New Zealand.
The New Zealand population of the Little Bittern is considered to be extinct. Data are not sufficient to determine this with certainty. A survey is needed to better determine whether the population persists.
Survey potential habitat of the Little Bittern in New Zealand to determine its persistence.
Australasia 5. Distribution and status of Sumatran Heron
Goal: Clarification of the current status and distribution of Sumatran Heron populations.
The Sumatran Heron is widespread but generally is reported to occur in only small numbers in any location. It uses lowland forests, a habitat under threat in most areas. It is widespread in distribution but sparse locally. It has been shown to abandon feeding and breeding areas close to human habitation. In New Guinea, considerable appropriate habitat remains and there seems to be no immediate threat in northern Australia, except perhaps along the developing east coast. Its status on Pacific islands is unclear. More information is needed on actual distribution and important sites. Its scarceness and solitary nature makes surveys and monitoring difficult, so information may need to be gleaned from numerous sources.
Survey status of Sumatran Heron over its range; identify important areas.
Australasia 6. Conservation of the Striated Heron in Chagos, Maldives and Diego Garcia
Goal: Protection of the Striated Heron in the Chagos, Maldives, and Diego Garcia.
The Striated Heron populations in Chagos, the Maldives, and Diego Garcia are considered to be vulnerable because of their restricted range on offshore islands. Populations on each island group were once considered separate subspecies (Butorides striatus albolimbatus, Butorides striatus didi, Butorides striatus albidulus), suggesting some level of distinctiveness. Status of the populations are not well known, and a survey is needed to determine population size and important sites on each island. Threats include the US military activities on Diego Garcia and increasing tourism on the Maldives. The status of the population on Chagos Islands is of particular interest, given the lack of human settlement there in the past decades. For all these islands, it is timely to develop a priority list of areas and sites for full protection.
Conduct survey of the Striated Heron in Chagos, the Maldives, and Diego Garcia; determine important sites for preservation; conduct a status and taxonomic review of the populations.
Australasia 7. Conservation of the Grey Heron in Sumatra
Goal: Assessment of the conservation status of the Grey Heron in Sumatra.
The Grey Heron population of Sumatra is much restricted and so is considered to be vulnerable. This population has previously been described as a distinct subspecies (Ardea cinerea altirostris). It is restricted to the island, where it occurs over a limited range of habitats, most of which are under some degree of threat. Information on waterbird populations on the island is sparse, so it is not currently possible to provide a population estimate greater than 700 mature individuals. Surveys covering the known or suspected distribution of the Grey Heron in Sumatra are needed to improve knowledge of taxonomic status, distribution and abundance. If the results suggest that there is a real cause for concern, then a more intensive conservation strategy should be prepared.
Conduct survey and assess conservation status and needs of the Grey Heron in Sumatra.
Australasia 8. Conservation of the Indian Pond Heron in the Maldives
Goal: Protection of the Indian Pond Heron in the Maldives.
The Indian Pond Heron population in the Maldives is vulnerable. It is highly restricted, exposed to pressure from tourism and habitat degradation. In the short-term, the most important action must be to determine the current size of the population, together with an assessment of habitat availability, and the degree of threat to breeding, and foraging habitats. The extent to which the species has a market in the Maldives needs to be determined and, if this exists, it needs to be controlled. Promotion of conservation awareness clearly needs to be a fundamental component of a conservation project.
Conduct an inventory of the population in the Maldives; determine and protect important sites; undertake a public awareness campaign to stop local capture and use of the species.
Australasia 9. Conservation of the Black Bittern in Australia
Goal: Clarification of conservation status, distribution, and needs of the Black Bittern in Australia.
The Black Bittern population in Australia is poorly known and suffering from wetland loss, habitat destruction and water problems. Despite lack of data, it is considered to be decreasing and is considered vulnerable overall in Australia. It has been classified as endangered in Victoria. It is necessary to establish its conservation status on a regional and national basis and to determine conservation strategies for its survival.
Determine conservation status of Black Bittern in Australia and ascertain conservation needs.
Australasia 10. Conservation of Little Bittern in Australia
Goal: Better determination of conservation status and needs of the Little Bittern in Australia.
The status of the Little Bittern is very poorly known in Australia, but it is uncommon and probably threatened by wetland loss and water management. It is thought to be decreasing and has been listed as a species of concern by two Australian states. It is necessary to determine its conservation status on a regional and national basis and to provide conservation strategies for its survival.
Determine conservation status of Little Bittern in Australia; ascertain conservation needs.
Australasia 11. Network of important heron sites in Australia
Goal: Establishment of a network of important sites for herons throughout Australia.
Many important sites for heron colonies are known in Australia. Some of these change from year to year, making preservation of particular sites more difficult. Based on available information and new inventories, colony sites used by herons throughout Australia should be identified and collated in a data base. These should be recognized as important sites for herons and networked to assist in their conservation. Given the vagaries of the Australian climate, these sites would not be expected to be used every year, but still should be protected to allow movement among sites in different years. This network could effectively be part of a more-encompassing waterbird site network.
Establish a network of sites important to herons around Australia.
America
Many species of herons are found throughout the Americas. In southerly locations both breeding and wintering populations intermingle seasonally.
Post breeding dispersal and migration result in large-scale annual movements of birds. For conservation purposes the entire New World is treated as a single region. The region includes North America, Central America, The Caribbean, and South America, as well as biogeographically (and politically) connected islands such as Bermuda, Falklands, Hawaii, and Galapagos.
South America, with its numerous and diverse wetlands and extensive floodplain forests, supports a particularly rich heron fauna, several genera and species being endemic to the continent. Data on nearly all populations are severely limited, but especially for species that nest singly or form small and dispersed colonies. Populations cannot now be estimated to any realistic degree of certainty, and even details of ranges are poorly documented. South America hosts some of the least understood herons of the world. To acquire more knowledge
is at present a major concern and also a challenge in view of the inaccessible areas to cover, such as the huge Amazon and Pantanal Basins. In Central America, only recently has information begun to be assembled. In the Caribbean, heron populations are small and limited habitats on various islands put populations at risk. The Caribbean, Central America and northern South America hold habitat of importance to migrating and wintering North American populations. In contrast to the Neotropics, heron populations have been rather extensively studied in North America. Important sites are known for most species, and many are protected. A continental nesting data base exists, but regional or national initiatives for coordinated long-term censuses of colonial herons have not succeeded to date. An extensive, cooperative, infrastructure for bird conservation exists in Canada, USA, and Mexico, in which herons are included. There is a continuous need for improving monitoring programs in order to gain insight into status and trends and for the protection of nesting sites.
America 1. Distribution and status survey of herons in the Neotropics
Goal: Better understanding status of herons in Latin America and the Caribbean.
Population sizes, and even the distribution, of species of herons in the Neotropics are very poorly known. With the exception of some local populations, nearly no information exists on numbers and important sites for herons within most countries. Areas of particular concern are the Amazon, Pantanal, Llanos, Central America and the islands of the Caribbean. Among species of particular concern are the Zigzag Heron, Agami Heron, tiger herons, Butorides herons, Cocoi Heron, Reddish Egret, Great Blue Heron (dimorphic form), Capped Heron, Whistling Heron, and the newly colonizing Little Egret. Information on all species in all nations of the region needs to be improved through national inventories of nesting and wintering populations. Inventories should be organized on a national basis and the results communicated to both national agencies and national environmental organizations as appropriate. Information from across the region should be collated and synthesized into regional and national assessments. Hemispheric partnerships among governmental and nongovernmental conservation organizations, researchers, and nature tour guides and other groups are a way to gain information. A hemispheric web-based data entry system should be developed. Among existing networks, sites important to herons need to be identified and joined in a hemispheric communication network.
Conduct national surveys of heron population sizes throughout the Neotropics; develop a hemispheric data base for population information.
America 2. Population sizes and trends of herons in North America
Goal: Better quantification of the population sizes and trends of herons in North America.
North America has a long and extensive record of censuses and surveys of waterbirds, including herons. Yet estimating continental population sizes remains difficult due to the complexity, incompleteness, and incompatibility of the existing record. Given the attention bird conservation, including herons, receive in North America, it is essential that existing data be better evaluated and that heron monitoring be more encompassing, more consistent, and better coordinated. The existing North American data base for waterbird censuses needs to be continued and enhanced. Evaluation of these existing records to better estimate populations of heron in North America is needed, with due consideration for comparability. A continental monitoring program is needed that is both sufficiently accurate and sufficiently precise to show population size and trends.
Determine population size, and trends of herons in North America; continue North American Colonial Bird data base; establish a continent-wide program to monitor nesting colonies.
America 3. Important sites for herons in North America
Goal: Identification and monitoring of important sites for herons in North America.
The identification of important areas for birds is well underway in North America, at both national and sub national scales. It is now or will soon be possible to extract from the overall list, a selection of sites important to herons. These sites should be identified, systematized, monitoring programs developed, and conservation needs assessed.
Evaluate existing information to determine most important sites for herons; develop monitoring and conservation programs at these sites.
America 4. Herons and conservation partnerships in the Americas
Goal: Incorporation of herons in programs emanating from more inclusive conservation initiatives.
In Canada, USA, and Mexico, considerable progress has been made in developing coordinated approaches to bird conservation. The North American Bird Conservation Initiative serves as a facilitating body for much of bird conservation in the three countries, and the Waterbird Conservation for the Americas initiative does the same for waterbirds throughout the hemisphere. It is essential that heron conservation continue to be a part of these programs. Public lands, states within the United States, and non-governmental conservation organizations have recognized their roles in waterbird conservation, and many are initiating waterbird conservation planning and action on the ground, especially colony site protection. Wetland managers have particularly begun to include heron conservation within their activities. Herons need to continue to be a concern within these programs. Heron conservationists should participate in multi-species bird conservation planning, programs, initiatives, and conservation activity assuring that the needs and opportunities for heron conservation are a part of these larger conservation programs. Parks, refuges, nature reserves, national forests, private reserves, wildlife management areas, and similar protected areas have been set aside throughout the Americas, especially in North America. Mangers of these reserves must be at the forefront of conserving herons on their site, and so should be encouraged to consider the needs of herons in their plans and activities. There is a need to provide funding and training for local conservation organizations and individuals that could be encouraged to adopt and protect colonies.
Incorporate heron conservation within more encompassing bird conservation initiatives and management of protected areas.
America 5. Status and distribution of the Zigzag Heron
Goal: Better understanding of the status of Zigzag Heron throughout South America.
The status and biology of Zigzag Heron is nearly unknown. It has been reported only from a few sites in the greater Amazon, although it is expected to occur throughout forested tropical South America. Recent observation extend its confirmed range into the Orinoco and Llanos. It is a highly unusual heron in its morphology and likely its habits, but its biology is little understood and needs to be studied. While it is not likely that surveys can be conducted over its potentially vast range, a program to encourage reports on its occurrence can improve information on its dispersion, habitat, and population centers. Any location where more than a few occur, at this point, should be considered an important site for the species.
Improve knowledge of the distribution and habitat use by the species through encouraging and synthesizing reports; protect all nesting sites discovered.
America 6. Status and biology of the Agami Heron
Goal: Better understanding status and nesting biology of the Agami Heron.
The Agami Heron is quite an unusual heron. Its biology is little known, as shown by the recent discovery that it sometimes nests in large colonies. Likely there is much to be learned about its ecology and behavior that is of value to its conservation. Needed investigations include breeding behavior, foraging dispersion during nesting and year-round movements. Its dependence on fragile river and riparian forests needs to be better understood. Concentration in large colonies raises the importance of identifying and protecting these colony sites. Large nesting sites should be sought, characterized, and protected. Sites presently known in Belize and French Guiana are top priorities. Feeding areas associated with nesting colonies
need to be determined.
Investigate biology and status of the Agami Heron; locate and protect large colony sites and associated feeding areas.
America 7. Distribution and status of Great White Heron and Reddish Egret in Caribbean
Goal: Clarification of the status of the Great White Heron and Reddish Egret on Caribbean coasts.
Two primarily North American herons are restricted to tropical coastal environments, which are degrading region wide. The conservation status of the dimorphic form of the Great Blue Heron, called the Great White Heron, and the Reddish Egret, also dimorphic, are of concern. These both may be declining in their North American strongholds. Records of individuals and small nesting populations of the Great White Heron and the Reddish Egret have been reported from several areas of the Caribbean and Central America. But the true status of these populations is not known. Given downward trends in the population core in North America, it is imperative to understand the full distribution of these herons.
Conduct surveys and establish communication networks to determine the status of Great White Heron and Reddish Egret along Caribbean coasts.
America 8. Conservation of the Reddish Egret in North America
Goal: Better delineation of the conservation status of the Reddish Egret in North America.
The Reddish Egret has a restricted geographical range and exploits a vulnerable coastal habitat. The total global population of the Reddish Egret is estimated to be about 12000-18000, although populations outside the United States may number fewer than 1000. Due to apparent declines, the population segments in North America are considered to be vulnerable. This dimorphic species is essentially restricted to shallow, tropical lagoons, which are suffering from ecological changes in recent years. Changing hydrology through building dikes and canals, thus altering the natural functioning of a coastal wetland, reduces the quality of the shallow flats on which these birds feed. Estuaries and coastal lagoons are exposed to contamination and under constant threat from development. Migratory pathways of North American Reddish Egrets are little known. There have been no recent population surveys. The genetics of the various populations are not understood. Important nesting sites need to be identified, and monitoring programs put in place. Fortunately, a Working Group has been established to coordinate research and conservation programs.
Survey the numbers of nesting Reddish Egrets in North America; determine populations of conservation concern; identify and protect important nesting sites and the related foraging habitat.
America 9. Conservation of the Great White Heron in Florida
Goal: Conservation of the Great White Heron in Florida.
The Great Blue Heron population found in the extreme south of southern Florida and in parts of the Caribbean and Central America is unusual for the high proportion of individuals having all white plumage. This population, called the Great White Heron, has previously been recognized as a distinctive species and is more recently considered to be a subspecies of the Great Blue Heron. The population clearly differs from continental Great Blue Herons, but the extent of that differentiation remains unclear despite repeated study. In South Florida the Great White Heron has a restricted range in coastal lagoons. The population in South Florida is thought to be about 1000 birds. A long term monitoring program in the southern Florida Keys show the population to be decreasing there. There has not been consistent monitoring elsewhere in its Florida range Fortunately nesting sites in Florida are protected. The cause of the decline in the Florida Keys is not known. In Florida Bay, the population has been shown to be under food stress. Little is known about seasonal movements, nesting success, survivorship, and year- round habitat use and needs. Causes of the apparent population decreases in Florida Bay and in the lower Florida Keys need to be determined quickly as the downward trend is rapid.
Conduct censuses of the Great White Heron in Florida; develop monitoring programs; further evaluate population structure and demography; determine and reverse causes of population declines.
America 10. Conservation of the Little Egret in Barbados
Goal: Protection of the Little Egret in Barbados.
The Little Egret recently colonized the Western Hemisphere as a nesting species, becoming established in Barbados, West Indies. Its population there has not significantly increased over the decade since its colonization. It nests on a single tiny site and depends for feeding on the very few remaining wetlands on the island. Because of its small population and restricted habitat, this population is considered to be extremely vulnerable. The single nesting site, in a private nature reserve, is degrading and requires active management before it disappears. Most feeding sites are not officially protected. In fact most sites are privately owned and managed and so have no assurance of persistence in long term. Current plans for parks in and near the present nesting sanctuary would decrease available habitat. Active management of the nesting site and protection of remaining wetlands, including privately maintained wetlands, are essential. The public in Barbados seems little aware of the presence of the species, and information needs to be provided to the public and the government. Individual birds are increasingly seen on other islands, suggesting dispersal, and the status of these birds needs to be better understood.
Manage nesting site of the Little Egret in Barbados; protect and enhance all wetlands used in Barbados; begin a public awareness campaign to highlight the species’ presence and conservation needs.
America 11. Conservation of Green Heron in Bermuda
Goal: Protection of the nesting sites and feeding areas of Green Heron in Bermuda.
The Green Heron recently colonized Bermuda and nests at only a few sites. Due to its recent colonization, few nesting sites, and small population, this population is considered to be vulnerable. It is recently nesting at more sites, and the population appears to be expanding. This presents a nearly unique opportunity to document
a natural colonization in detail. The changing status of the population should be documented thoroughly. All nesting sites need to be identified, and all nesting sites and feeding sites need to be protected. This likely involves conserving all of the remaining mangroves and freshwater wetlands on the island. Wetlands are so few, that creation of additional wetlands would benefit herons, and other species, by providing additional foraging and nesting opportunities. Among its prey, the heron appears to eat an endemic killifish. The potential impact of the heron on the fish needs to be understood and if a problem exists, ways found to accommodate both species. Popularization of the presence of the Green Heron, as a new species to Bermuda, would provide public support for conservation of its habitat.
Conduct surveys to document the changing status of the species on Bermuda; protect all nesting and feeding sites; better popularize the species.
America 12. Population status of the Yellow-crowned Night Heron on Bermuda
Goal: Re-assessment of the population status of the Yellow-crowned Night Heron on Bermuda.
The Yellow-crowned Night Heron was purposely introduced into Bermuda. It is considered to be replacing an extinct form of the night heron. It was introduced to control land crabs, and the experiment succeeded. The species is now well established on the island. It is time to reassess the population size, and synthesize and further evaluate the history of the introduction. It is also important to document its important nesting sites to assure their future conservation through island-wide conservation planning.
Reassess status of the Yellow-crowned Night Heron in Bermuda; identify important sites; develop conservation plan for protecting these sites.
America 13. Status of the Least Bittern in Peru
Goal: Understanding status and distribution of Least Bitterns in Peru.
The Peruvian populations of the Least Bittern are little understood. Ixobrychus exilis peruvianus is an apparently disjunct population in several lagoons and marshes along a narrow coastal belt of Central Peru, a heavily populated area, Ixobrychus exilis erythromelas occurs in the north and east of the country. Its status and distribution is not sufficiently understood. Fishing, duck hunting, reed cutting for basket making, pesticide run-off from adjacent agriculture, public recreation, livestock grazing and drainage likely affect wetlands throughout this range. To better understand the status of the species, an inventory is needed and important sites determined throughout Peru. Also the subspecific systematics of these populations needs to be better studied.
Conduct nationwide inventory; determine important sites; investigate taxonomic distinctiveness of Least Bitterns in Peru.
America 14. Status and conservation of American Bittern in North America
Goal: Better documentation of the status, important sites, and trends for American Bitterns throughout North America and development of conservation actions at important sites.
The nesting sites of the American Bittern in North America are poorly documented. Difficulties in techniques have inhibited development of a census and monitoring program, so neither population size nor trend is well understood. It is thought, however, that the population is decreasing and its range retreating northward. Recent developments in acoustic sampling, in both Europe and America, hold promise that the species finally can be appropriately censused and monitored. It is necessary to fine tune the protocol and put it into place to determine range, population size, and important areas for American Bitterns throughout North America. Most sites are likely on protected areas in North America, and managers of these sites should be enlisted to conduct the monitoring and to develop conservation plans at each site. The causes of the apparent decrease in the population and range are not known, but it is similar to the situation faced by the other great bittern species. Determination of the reality and cause of decline is needed.
Adopt a standardized census protocol and conduct surveys of the American Bittern throughout its breeding range; increase investigations of the cause of the apparent population decrease; identify, protect, and manage important nesting areas.
America 15. Status of the Streaked Bittern in South America
Goal: Better understanding of the status and distribution of the Streaked Bittern.
The status of the Streaked Bittern is not well understood. It has been thought to be divided into two populations, one in northern South America and one in southern South America. But recent records from between these ranges suggest it may be more widespread. The range needs to be clarified. A third apparently disjunct population is found on the coast of Chile. There is almost no information on this Chilean population. It has been reported from Aconcagua, Region Metripolitana de Santiago, Libertador General Bernado O’Higgins, Maule, Bio-Bio, La Araucana and Los Lagos provinces, but the presence and status of herons in this area remain unclear. Habitat requirements of the several populations are unclear, given its wide range. Collection of birds and eggs for subsistence and trade may have a significant effect. Determining it range, conservation issues, and patterns of geographic variation are high priorities.
Conduct a survey of the status, distribution and geographic variation of the Streaked Bittern in South America.
America 16. Status and biology of the Bare-throated Tiger Heron
Goal: Better understanding of the status and biology of Bare-throated Tiger Heron.
The Bare-throated Tiger Heron is the only heron whose population is centered in Central American. It generally has been thought to occur only along streams and forests. However on the Pacific coast, it has recently been documented to also use rocky shorelines, mangroves swamps, and even beaches. Clearly there is much to be understood about habitat use, distribution, and biology of this species throughout its range. Status of the species in the Gulf of Panama, coastal Mexico, and extreme northern Colombia needs to be better determined.
Determine the distribution, conservation status, and taxonomic status of all populations of the Bare-throated Tiger Heron; better delineate habitat use.
America 17. Status of the Fasciated Tiger Heron in the Neotropics
Goal: Better understanding of the status of the Fasciated Tiger Heron in the Neotropics.
The Fasciated Tiger Heron occurs in South and Central America, although the details of its range are not clear. Populations appear to be separated by the Andes and Amazon, and a third subspecies is recognized from Argentina. Recent records from northeastern South America suggest a wider range. Patterns of geographic variation need to be better studied, using molecular techniques. It is a very little known species biologically. It seems to occupy stream edge forests, a vulnerable habitat. Its populations are likely small, and likely threatened by habitat loss. Its range needs to be better understood and important areas identified.
Evaluate distribution, population size, and geographic variation of the Fasciated Tiger Heron throughout its range.
America 18. Systematics of Butorides Herons
Goal: Better definition of systematic relationships among Butorides populations.
Presently, the Striated Heron and Green Heron are considered to be separate species, although they previously had been considered the same, as had the population on the Galapagos. Different studies of specimens come to different conclusions, suggesting the need for use of additional characters, such as those molecular studies can provide. It is likely that the two forms interbreed in the southern Caribbean, although there is no evidence of interbreeding where their ranges meet in Central America. Several subspecies of Green Heron have described as has the distinctive population on the Galapagos. Study of the taxonomy of Butorides in the America using molecular techniques is needed. Expanding the study worldwide would be even more definitive. Knowing the patterns of geographic variation and evolutionary history of the group is needed to determine conservation strategy for the populations.
Re-examine the geographic variation of Butorides populations in the Americas.
America 19. Systematics of the Boat-billed Heron
Goal: Clarification of the taxonomy of the Boat-billed Heron.
It appears that the Boat-billed Heron is part of an ancient linage of herons. Several populations have been recognized as subspecies based on plumage differences. It is likely that the patterns of geographic variation and taxonomy of the Boat-billed Heron are not yet sufficiently clarified. There may be other subspecies or even species within this complex. Molecular techniques have not been applied to the species/subspecies taxonomy of this intriguing species. Knowing the evolutionary relationships within the taxon is important to defining conservation objectives for populations.
Conduct taxonomic study of Boat-billed Herons throughout its range, using molecular techniques.
America 20. Status and distribution of the Capped Heron
Goal: Better understand the population status, distribution, and biology of the Capped Heron.
The Capped Heron, a distinctive neotropical heron, is poorly known. It is thought to be an Amazonian species, but records of the details of its distribution are few. Its biology of nearly unknown. Basic information on biology and distribution is needed.
Determine the distribution and population status of the Capped Heron.
America 21. Status of the Yellow-crowned Night Heron
Goal: Better understanding of the status of Yellow-crowned Night Heron populations throughout its range.
The Yellow-crowned Night Heron is a species widespread in North America, Caribbean and coastal Neotropics. Several populations have been recognized as subspecies, but other populations may also be distinctive. Because conservation most effectively takes place on a population basis, the status of the heron in its various populations needs to be determined and conservation measures enacted as appropriate. Populations requiring particular attention include: Western Mexico; Central America; Panama; Tobago – especially Kilgwyn Swamp, Bon Accord Swamp, and Boccoo Reef reserve; coastal Colombia; Galapagos (the most distant population); Bermuda; and Caribbean Islands.
Conduct status surveys and study patterns of geographic variation of Yellow-crowned Night Heron populations in various parts of its range.
America 22. Status of the Little Blue Heron
Goal: Better understanding of the conservation status of the Little Blue Heron throughout its range.
The Little Blue Heron is considered to be of conservation concern in North America. Due to difficulty in censusing this species, concern is based primarily upon a lack of information. Its status is similarly little known through the rest of its range. In order to determine the conservation needs of this species, its true conservation status needs to be determined in North America and also through the rest of its extensive range. A focused survey is needed to determine nesting populations in North America. A general idea of status also needs to be determined in the rest of its range, likely thorough development of a communication network.
Conduct a conservation status assessment of the Little Blue Heron in North America and determine overall range-wide status.
America 23. Conservation of the Great Blue Heron in Western North America
Goal: Establishment of a network of protected nesting and feeding sites for the Great Blue Heron in northwestern United States and southwestern Canada.
The Great Blue Heron population of the coastal Washington, USA, and British Columbia, Canada, is recognized as distinct subspecies, a determination that recent study reconfirmed. The population is restricted and becoming fragmented. Its reproductive success has been declining in recent years as nesting colonies are increasingly predated by Bald Eagles. The population is about 10000 individuals, so at times remains robust. There is a need to establish a network of protected foraging and nesting sites that qualify as important sites for the subspecies. Numbers and reproductive success need to continue to be monitored. Sufficient habitat should be secured to ensure a viable population of the Great Blue Heron on the coast of Washington and British Columbia.
Determine and establish a protective network of important sites for nesting and foraging of the Great Blue Herons in coastal Washington and British Columbia.
America 24. Status and biology of the Lava Heron in the Galapagos
Goal: Better understanding of the status and biology of the Striated Heron population on the Galapagos Islands.
The Striated Heron of the Galapagos Islands, called the Lava Heron, is distinctive in both its plumage polymorphism and biology. Although widespread, on the shores of the islands, its biology is little reported. Nor is its status and population size well understood. The population deserves increased attention for both its biology and conservation. A monitoring program should be put in place to assure its continued status.
Better understand biology and status of Striated Herons on the Galapagos; establish monitoring program.
America 25. Status of Old World herons in the western hemisphere
Goal: Better understand status of Old World Herons in the western hemisphere.
The Grey Heron, Little Egret, and Western Reef Heron are observed with some regularity in the Western Hemisphere, particularly in the West Indies, where the Little Egret has become established on Barbados. It is being seen on other islands in recent years. Recording the Grey Heron and white form of the Little Egrets, superficially similar to North American herons, require observers be aware of their potential presence. The dark form of the Little Egret (also known as the western Reef Heron) has been observed along the North American east coast. The possibility of these species being more common than presently thought needs to be examined so that occurrence and possible colonization of the hemisphere can be documented.
Increase awareness and reporting of Grey Herons, Reef Herons and Little Egret sightings in the Western Hemisphere, especially in the West Indies.
America 26. Conservation of herons in the Everglades
Goal: Restore populations of herons in the Everglades of southern Florida.
The extensive marshes of the Florida Everglades were once one of the key areas for herons in North America. These marshes historically supported all North American species. Nearly all have nesting populations, and all winter in the area. Populations of distinctive forms such as the Great White Heron and Reddish Egret depend on coastal zones supplied by runoff from the Everglades. All evidence suggests that heron populations have declined since the institution of management of water in the Everglades, especially in the past 40 years. Much planning and resources are being devoted to restoring the Everglades. Within this program, the restoration of heron populations needs to be a high priority. Monitoring and restoration focus recently has been on a couple species that are readily counted. All species need attention, however.
Restore heron populations in the Everglades.
America 27. Reduce conflicts between herons and aquaculture
Goal: Institution of policies to reduce conflicts between herons and aquaculture throughout hemisphere.
In many areas of the Americas, herons come into conflict with aquaculture facilities. Thousands of birds are killed yearly at aquaculture facilities in the United States, even where laws are in effect that can regulate this killing. In other parts of the hemisphere legal protections are not present or enforced. Killing herons can result in local reduction of populations, but seldom effect levels of depredation. Killing herons outside the breeding season can effect populations over much larger areas than the facility itself. Studies are needed to determine the true impact of depredation at various types of facilities. It should be encouraged to design facilities that do not encourage predation by herons. Laws, regulations, and regulatory policies need to assure that local populations of herons are not adversely affected by heron control at aquaculture sites.
Reduce impact of control of herons at aquaculture facilities.
References
BirdLife International. 2004. Threatened Birds of the World 2004. BirdLife International.
Delany, S. and D. Scott. 2006. Waterbird Population Estimates. Fourth Edition. Wetlands International. Wetlands International, Wageningen, Netherlands.
Hafner, H. and Kushlan, J.A. 1990. ICBP/SSC Heron Specialist Group. Species 15:79.
Hafner, H. and Kushlan, J.A. 1996. WI/BirdLife/SSC Heron Specialist Group. Species 26-27: 127.
Hafner, H. and Kushlan, J. A. (Eds.). 2002. Action plan for Conservation of the Herons of the World. Heron Specialist Group. Station Biologique Tour du Valat, Arles, France.
Hafner, H., Dugan, P., and Kushlan, J. A. 1986. The ICBP/IWRB Herons Specialists Group: Origin, present and future. Colonial Waterbirds 9:126-127.
Hafner, H. R.V. Lansdown, J.A. Kushlan, and R. Butler. 1996. Action Plan for the Conservation of the Ardeidae. Herons Specialist Group and International Waterbird and Wetlands Research Bureau, Slimbridge UK.
Kushlan, J. A. and Hafner, H. 1991. ICBP/IWRB/SSC Heron Specialist Group. Species 16:72.
Kushlan, J.A. and Hafner, H. 1993. An evaluation of the status and conservation of the herons of the world. Species 19:68.
Kushlan, J.A. and Hafner, H. 2000. Heron Conservation. Academic Press, London and San Diego.
Kushlan, J.A. and Hancock, J.A. 2005. The Herons. Oxford University Press, Oxford.
Acknowledgements
Many people and institutions have contributed information used in assessing the status and proposing conservation actions for herons. The members of the Heron Specialist Group have provided much information for this report. Individuals providing information and suggesting changes from the previous editions of the Plan include: Neil Baker, Bruce Beehler, Stuart Butchart, Robert Butler, Bosco Chan, Xiaolin Chen, Malcolm Coulter, Mike Crosby, Simon Delany, Robert Dickerman, Tim Dodman, Zhou Fang, Mauro Fasola, Jack Fellowes, Jacques Franchimont, Stephen Garnett, P. Gerlach, Heinz Hafner, James Hancock, Doug Harebottle, Chuck Hunter, Roger Jaensch, Cathy King, H. Van der Kooij, Richard Lansdown, David Li, Yu Lijiang, Max Maddock, Stefani Melvin, Nial Moores, Gonzalo Morales, Taej Mundkur, Tshewang Norbu, Fabio Olmos, Marc Rabenandrasana, Hugo Rainey, Mark Riegner, Jack Tordoff, Christophe Tourenq, Hishey Tshering, Don Turner, Stephanie J. Tyler, Stefano Volponi, Thomas Wilmers, Duncan Wilson, Captain L. C. Wong, H. Glyn Young, and Lew Young. We thank Patrick Grillas, Coralie Hermeloup, Marc Thibault and James Hancock for their many contributions to the publication of this report.
The Heron Specialist Group acknowledges the decades of support and encouragement received from our sponsoring organizations, The International Conservation Union Species Survival Commission, Wetlands International, and its predecessor the International Waterbird and Wetland Research Bureau, as well as our other long term institutional partners BirdLife International and Station Biologique de la Tour du Valat.
Breeding Range (Non-breeding Range in parentheses)
Tigriornis leucolopha
White-crested Tiger Heron
AF : Senegal to Central African Republic and DR Congo
Tigrisoma lineatum
T. l. lineatum Honduras s to ne Bolivia and Amazonian Brazil
T. l. marmoratum se Bolivia to s Brazil and n Argentina
Rufescent Tiger Heron
MA, SA : widespread
Tigrisoma mexicanum
Bare-throated Tiger Heron
MA, SA : Mexico to Colombia
Tigrisoma fasciatum
T. f. salmoni Costa Rica to n Bolivia
T. f. fasciatum se Brazil to n Argentina
T. f. pallescens nw Argentina
Fasciated Tiger Heron
MA, SA : Costa Rica to n Bolivia and se SA
Cochlearius cochlearius
C. c. zeledoni w Mexico
C. c. phillipsi e Mexico and Belize
C. c. ridgwayi s Mexico to Honduras
C. c. panamensis Costa Rica and Panama
C. c. cochlearius n, c South America
Boat-billed Heron
MA, SA : wc Mexico to ne Argentina
Agamia agami
Agami Heron
MA, SA : e Mexico to Amazonia
Zebrilus undulatus
Zigzag Heron
SA : Amazonia
Botaurus stellaris
B. s. stellaris Europe to e Asia
B. s. capensis s Africa
Eurasian Bittern
PAL, AF : widespread
Botaurus poiciloptilus
Australasian Bittern
AU : far sw WA, se QLD to se SA and Tasmania (sw, se Australia), North, South and Stewart is. (New Zealand), Ouvea (Loyalty Is.) and Grande Terre (New Caledonia; rare)
Botaurus lentiginosus
American Bittern
NA : widespread
Botaurus pinnatus
B. p. caribaeus se Mexico and Belize
B. p. pinnatus El Salvador and Nicaragua to n Argentina and s Brazil
Pinnated Bittern
MA, SA : e Mexico to ne Argentina
Botaurus involucris
Stripe-backed Bittern
SA : Colombia to Suriname, s Bolivia and s Brazil to c Chile and c Argentina
Botaurus exilis
B. e. exilis e Canada and e, sw USA
B. e. pullus nw Mexico
B. e. erythromelas e Panama and n South America to n Bolivia and n Argentina
B. e. limoncochae e Ecuador
B. e. bogotensis c Colombia
B. e. peruvianus wc Peru
Least Bittern
NA, MA, SA : e, sw USA to se Brazil
Botaurus flavicollis
B. f. flavicollis India to Indochina, Philippines and w, c Indonesia
B. f. australis e Indonesia, New Guinea, Bismarck Arch. and Australia
B. f. woodfordi Bougainville to Rennell (n to s Solomon Is., except Makira)
Black Bittern
OR, AU : widespread
Botaurus cinnamomeus
Cinnamon Bittern
OR, PAL : Pakistan and Maldives through se, e Asia to e China, Ryukyu Is. (s Japan), s to Philippines and w, c, Indonesian Arch.
Botaurus eurhythmus
Von Schrenck's Bittern
PAL : se Siberia, Korean Pen., Japan and e China
Botaurus sturmii
Dwarf Bittern
AF : widespread
Botaurus minutus
B. m. minutus c, s Europe to c Asia and nw India
B. m. payesii Africa s of the Sahara
B. m. podiceps Madagascar
Little Bittern
PAL, AF : s, c Europe to c Asia, nw India and Africa
Botaurus sinensis
Yellow Bittern
PAL, IO, OR : Oman, Inner Is. (ne Seychelles), Pakistan and Sri Lanka through se Russia and Japan s to se Asia, Indonesian Arch., Philippines to w Micronesia
Botaurus dubius
Black-backed Bittern
AU : patchily in sc, nc, se New Guinea and far sw WA, ne WA to s VIC (sw, n, e Australia)
Botaurus novaezelandiae
New Zealand Bittern
AU : South I. (New Zealand)
Nyctanassa violacea
N. v. violacea c, e USA to e Mexico and e Costa Rica
N. v. bancrofti w Mexico to w Nicaragua, Socorro I. and West Indies
N. v. caliginis Panama and w Colombia to Peru
N. v. cayennensis Panama and ne Colombia to ne Brazil
N. v. pauper Galápagos
Yellow-crowned Night Heron
NA, MA, SA : e USA to Peru and e Brazil
Nyctanassa carcinocatactes
Bermuda Night Heron
AO : Bermuda
Nycticorax nycticorax
N. n. nycticorax Eurasia s to Africa and Madagascar and e to e Asia, Philippines and Indonesian Arch.
N. n. hoactli s Canada to n Argentina and Chile; Hawaii
N. n. obscurus Chile and sw Argentina
N. n. falklandicus Falkland Is.
Black-crowned Night Heron
Worldwide : except AU
Nycticorax olsoni
Ascension Night Heron
AO : Ascension I.
Nycticorax duboisi
Reunion Night Heron
IO : Réunion (w Mascarenes)
Nycticorax mauritianus
Mauritius Night Heron
IO : Mauritius (c Mascarenes)
Nycticorax megacephalus
Rodrigues Night Heron
IO : Rodrigues (e Mascarenes)
Nycticorax caledonicus
N. c. crassirostris Bonin Is. (=Ogasawara Is.; se Japan)
N. c. manillensis Philippines and n Borneo
N. c. australasiae Java e to New Guinea, nw Bismarck Arch., Australia and New Zealand
N. c. mandibularis e Bismarck Arch. and Solomon Is. except Temotu (=Santa Cruz Is.; se Solomon Is.)
N. c. pelewensis Palau and Caroline Is. (s Micronesia)
N. c. caledonicus Grande Terre (New Caledonia)
Nankeen Night Heron
AU : widespread
Gorsachius melanolophus
Malayan Night Heron
OR : W Ghats (sw India), ne India through se Asia and se China to Philippines, Peleng (Banggai Is., e of Sulawesi)
Gorsachius goisagi
Japanese Night Heron
PAL : Honshu, Shikoku, Kyushu (c, s Japan), Jeju (off s Korean Pen.) and Taiwan
Pilherodius pileatus
Capped Heron
MA, SA : e Panama to se Brazil
Syrigma sibilatrix
S. s. fostersmithi ne Colombia and nw Venezuela
S. s. sibilatrix e Bolivia and s Brazil to ne Argentina
Whistling Heron
SA : n, sc, se
Egretta caerulea
Little Blue Heron
NA, MA, SA : se USA to s Brazil
Egretta tricolor
E. t. ruficollis e, se USA and e, w Mexico to nw South America and West Indies
E. t. tricolor nc, ne South America
Tricolored Heron
NA, MA, SA : se USA to n SA
Egretta rufescens
E. r. rufescens s USA, West Indies and Mexico
E. r. dickeyi Baja California (Mexico)
Reddish Egret
NA, MA : s USA and Caribbean
Egretta vinaceigula
Slaty Egret
AF : Zambia to ne Namibia, n Botswana and w Zimbabwe
Egretta ardesiaca
Black Heron
AF : widespread s of the Sahara
Egretta sacra
E. s. sacra coastal s Bangladesh and Andaman Is. to Ryukyu Is. (s Japan), Australasia (except s outlying is.), Melanesia (except New Caledonia) to Tuamotu Arch. (e Polynesia)
E. s. albolineata New Caledonia including Loyalty Is.
Pacific Reef Heron
OR, AU : coastal s Bangladesh and Andaman Is. to Ryukyu Is. (s Japan), Australasia (except s outlying is.), Melanesia to Tuamotu Arch. (e Polynesia)
Egretta eulophotes
Chinese Egret
PAL : coastal Russian Far East and Korean Pen. to e China
Egretta thula
E. t. brewsteri w North America
E. t. thula e, s North America to c Chile and n Argentina
Snowy Egret
NA, MA, SA : widespread
Egretta garzetta
E. g. garzetta Europe to Japan s to Africa, India and Mindanao (s Philippines)
E. g. nigripes Sundas to Australia and New Zealand
Little Egret
PAL, AF, OR, AU : widespread
Egretta dimorpha
Dimorphic Egret
AF : e, Europa (w of s Madagascar, Mozambique Channel), Aldabra group (sw Seychelles) and Madagascar
Egretta gularis
E. g. gularis coastal w Africa
E. g. schistacea coastal ne Africa to w India and Sri Lanka
Western Reef Heron
AF, PAL, OR : w AF, ne AF to w India and Sri Lanka
Egretta picata
Pied Heron
AU : Sulawesi and ne WA to ne QLD (n Australia)
Egretta novaehollandiae
White-faced Heron
AU : Timor (e Lesser Sundas), Australasia to New Caledonia, Fiji and Tonga (sw, sc Polynesia)
Calherodius leuconotus
White-backed Night Heron
AF : widespread
Oroanassa magnifica
White-eared Night Heron
OR : s, e China and n Vietnam
Butorides atricapilla
B. a. atricapilla Africa s of the Sahara
B. a. brevipes Somalia and the Red Sea coasts
B. a. crawfordi Aldabra and Amirante groups (s, c Seychelles)
B. a. rhizophorae Comoros
B. a. rutenbergi Madagascar and Réunion I.
B. a. degens ne Seychelles
B. a. albolimbata Chagos Arch. and Maldives
B. a. amurensis se Siberia, ne China and Japan
B. a. actophila e China to n Myanmar and n Vietnam
B. a. javanica Pakistan, India and Sri Lanka to Thailand, Philippines, the Greater Sundas and Sulawesi
B. a. spodiogaster Andaman and Nicobar Is. and islands off w Sumatra
B. a. steini Lesser Sundas
B. a. moluccarum Moluccas
B. a. papuensis nw New Guinea
B. a. idenburgi n New Guinea
B. a. flyensis sc, se New Guinea
B. a. stagnatilis coastal nw, nc Australia
B. a. macrorhyncha e, ne Australia and New Caledonia
B. a. solomonensis Lavongai (=New Hanover) to Solomon Is. (except Rennell), and Vanuatu to Fiji (sw Polynesia)
B. a. patruelis Tahiti Is. (Society Is.)
Little Heron
PAL, AF, AU : widespread
Butorides striata
Striated Heron
MA, SA: e Panama to n Argentina, Bolivia and Chile
Butorides sundevalli
Lava Heron
SA : Galápagos
Butorides virescens
B. v. virescens c, e USA and e Canada to Panama and the Caribbean
B. v. bahamensis Bahamas
B. v. anthonyi w USA and n Baja California (Mexico)
B. v. frazari s Baja California (Mexico)
Green Heron
NA, MA : e, c USA to Panama
Zonerodius heliosylus
Forest Bittern
AU : Salawati (Raja Ampat Is., nw of New Guinea), Aru Is. (sw of New Guinea) and New Guinea
Ardeola rufiventris
Rufous-bellied Heron
AF : Uganda and s Kenya to s Angola, n Botswana and e South Africa
Ardeola ralloides
Squacco Heron
PAL, AF : widespread
Ardeola idae
Malagasy Pond Heron
AF : Madagascar and Aldabra I.
Ardeola grayii
Indian Pond Heron
PAL, OR : Persian Gulf to Myanmar and Maldives
Ardeola bacchus
Chinese Pond Heron
PAL, OR : Assam (ne India) and n Myanmar through Russian Far East, Japan and s China
Ardeola speciosa
A. s. continentalis c Thailand through s Indochina
A. s. speciosa Sundas (w, c Indonesia) and Philippines
Javan Pond Heron
OR : Thailand through Philippines, Sulawesi region and Lesser Sundas
Ardea pacifica
White-necked Heron
AU : Australia
Ardea alba
A. a. alba c Europe to ne Asia s to n, c Africa and s Asia
A. a. melanorhynchos Africa s of the Sahara
A. a. modesta s, e Asia to Mindanao (s Philippines), Indonesian Arch. and Australasia
A. a. egretta s Canada to s Argentina and s Chile
Great Egret
NA, SA, AF, PAL, OR, AU : worldwide
Ardea brachyrhyncha
Yellow-billed Egret
AF : Africa s of the Sahara
Ardea intermedia
Medium Egret
PAL, OR: India to Japan and Greater Sundas
Ardea plumifera
Plumed Egret
OR: e Indonesia, New Guinea and Australia
Ardea ibis
Western Cattle Egret
PAL, AF, NA, MA, SA : s Europe to Iran, Africa, Indian Ocean Is., North to South America
Ardea coromanda
Eastern Cattle Egret
OR : s, e Asia and Australasia
Ardea cinerea
A. c. cinerea w Europe to e Asia to India and Africa
A. c. jouyi n China, Korean Pen. and Japan to Sumatra and Java
A. c. monicae Banc d'Arguin (Mauritania)
A. c. firasa Madagascar, Comoros and Aldabra
Grey Heron
PAL, AF, OR : widespread
Ardea herodias
A. h. fannini nw North America
A. h. wardi w, sc, s USA to n Florida and w, ne Mexico
A. h. herodias s Canada to nc, e USA s to the Carolinas
A. h. cognata Galápagos
A. h. occidentalis s Florida (USA) through the West Indies
Great Blue Heron
NA, MA : widespread
Ardea cocoi
Cocoi Heron
SA : widespread
Ardea purpurea
A. p. bournei Cape Verde Is.
A. p. purpurea s and c Europe to c Asia and the Middle East, Africa s of the Sahara
A. p. madagascariensis Madagascar
A. p. manilensis s, e Asia to Philippines and Indonesia
Purple Heron
PAL, OR, AF : widespread
Ardea humbloti
Humblot's Heron
AF : w, c Madagascar
Ardea insignis
White-bellied Heron
OR : e Himalayas
Ardea sumatrana
Great-billed Heron
OR, AU : se Asia and w Philippines to n Australia and New Guinea
Ardea melanocephala
Black-headed Heron
AF : widespread s of the Sahara
Ardea goliath
Goliath Heron
AF, PAL : widespread in Africa s of the Sahara, Iran and Iraq to India
Below are the latest estimates of the numbers of heron in various populations. Depending on the species, estimates may be for the entire species, a subspecies, or a geographic population. These estimates were derived by Wetlands International in partnership with HeronConservation and others. The entire report with information on the processes used may be obtained through the search page of Wetlands International.
Estimates are in all cases only that, and base on the best information available and best professional judgment of those who know the populations best. As information increases, estimates will be updated. HeronConservation will continue to partner with Wetlands International and the Ramsar Convention in periodically updating these estimates. All are invited to participate in making these estimates more accurate. Please communicate your data and suggestions to the HeronConservation Co-chairs or Steering Committee Members.
Heron Population Estimates
Scientific name
English name
Population
Range
Size - Year
Size
Trend - Year
Trend
Public-ation
Note
Zonerodius heliosylus
Forest Bittern
New Guinea
2006 - 2006
1 - 10,000
1985 - 2005
Declining
WPE5
References
Unpublished information supplied by Wetlands International Specialist Groups, 2006.
Notes
Very difficult to detect and invariably overlooked; thus virtually unknown in the wild (Bishop 2003).
Tigriornis leucolopha
White-crested Tiger-heron
W & C Africa
2006 - 2006
25,000 - 100,000
—
Unknown
WPE5
References
Unpublished information supplied by Wetlands International Specialist Groups, 2006.
Tigrisoma lineatum
Rufescent Tiger-heron
lineatum
2005 - 2005
2,700 - 5,500
—
Unknown
WPE5
References
Komar, O., Angehr, G., Eisermann, K., Herrera, N. and Zolotoff, J.M.. 2006. Waterbird Population Estimates for Central America. IV North American Ornithological Conference. unpublished report.
Notes
Estimate for whole of Central America.
marmoratum
2005 - 2011
25,000 - 100,000
2001 - 2010
Stable
WPE5
References
Barrett R. T. 1997. Br?nnich?s Guillemot Uria lomvia. Pp 370 in: Hagemeijer E. J. M and Blair M. J. (Eds) 1997. The EBCC Atlas of European Breeding Birds: Their Distribution and Abundance. T and A D Poyser, London.
Tigrisoma fasciatum
Fasciated Tiger-heron
fasciatum
—
1 - 10,000
—
Unknown
WPE5
References
Antas, P.T.Z. In litt. 1993. Unpublished data concerning South American waterbirds.
salmoni
2005 - 2005
2,520 - 5,050
—
Unknown
WPE5
References
Komar, O., Angehr, G., Eisermann, K., Herrera, N. and Zolotoff, J.M.. 2006. Waterbird Population Estimates for Central America. IV North American Ornithological Conference. unpublished report.
Notes
Includes Tigrisoma fasciatum, bolivianum.
Estimate for whole of Central America.
pallescens
2004 - 2004
2,000 - 2,000
—
Unknown
WPE5
References
Boyla, K. and Estrada, A. (Eds.) 2005. ?reas Importantes para la Conservaci?n de las Aves en los Andes Tropicales: Sitios prioritarios para la conservaci?n de la biodiversidad. Quito, Ecuador: BirdLife International and Conservation International (BirdLife Conservation Series No. 14).
Tigrisoma mexicanum
Bare-throated Tiger-heron
Colombia to Mexico
2005 - 2005
17,100 - 47,500
2005 - 2005
Increasing
WPE5
References
Komar, O., Angehr, G., Eisermann, K., Herrera, N. and Zolotoff, J.M.. 2006. Waterbird Population Estimates for Central America. IV North American Ornithological Conference. unpublished report.
S Mexico
—
—
—
Unknown
WPE5
Notes
Sometimes ascribed to fremitus.
Agamia agami
Agami Heron
C & S America
2005 - 2011
10,000 - 25,000
—
Unknown
WPE5
References
Komar, O., Angehr, G., Eisermann, K., Herrera, N. and Zolotoff, J.M.. 2006. Waterbird Population Estimates for Central America. IV North American Ornithological Conference. unpublished report.
Wetlands International Argentina (compiler). 2012. Unpublished information from the Neotropical Waterbird Census, 1990-2011, including contributions from Colombia (Asociaci?n Calidris in litt.), Paraguay (Guyra Paraguay in litt.), Uruguay (Averaves in litt.) and Venezuela (Uni?n Venezolana de Ornit?logos in litt.).
Cochlearius cochlearius
Boat-billed Heron
cochlearia
2011 - 2011
25,000 - 100,000
2001 - 2010
Declining
WPE5
References
Boyla, K. and Estrada, A. (Eds.) 2005. ?reas Importantes para la Conservaci?n de las Aves en los Andes Tropicales: Sitios prioritarios para la conservaci?n de la biodiversidad. Quito, Ecuador: BirdLife International and Conservation International (BirdLife Conservation Series No. 14).
Wetlands International Argentina (compiler). 2012. Unpublished information from the Neotropical Waterbird Census, 1990-2011, including contributions from Colombia (Asociaci?n Calidris in litt.), Paraguay (Guyra Paraguay in litt.), Uruguay (Averaves in litt.) and Venezuela (Uni?n Venezolana de Ornit?logos in litt.).
zeledoni
—
—
—
Unknown
WPE5
phillipsi
2005 - 2005
14,800 - 46,300
—
Unknown
WPE5
References
Komar, O., Angehr, G., Eisermann, K., Herrera, N. and Zolotoff, J.M.. 2006. Waterbird Population Estimates for Central America. IV North American Ornithological Conference. unpublished report.
Notes
Estimate for whole of Central America.
ridgwayi
—
—
—
Unknown
WPE5
panamensis
—
—
—
Unknown
WPE5
Zebrilus undulatus
Zigzag Heron
South America
2004 - 2004
1 - 10,000
—
Unknown
WPE5
References
Boyla, K. and Estrada, A. (Eds.) 2005. ?reas Importantes para la Conservaci?n de las Aves en los Andes Tropicales: Sitios prioritarios para la conservaci?n de la biodiversidad. Quito, Ecuador: BirdLife International and Conservation International (BirdLife Conservation Series No. 14).
Botaurus stellaris
Eurasian Bittern
stellaris
W Europe, NW Africa (bre)
2005 - 2012
7,150 - 9,100
2000 - 2012
Increasing
CSR7
References
BirdLife International 2015. European Red List of Birds. Luxembourg: Office for Official Publications of the European Communities. http://datazone.birdlife.org/info/euroredlist
Notes
In WPE2 this population belonged to one single population (Europe (breeding)).
2,375 - 3,013 pairs in BE, DE, DK, ES, FR, NL, PT, SE & UK (BirdLife International 2015). Less than 20 pairs in NW Africa (Dodman, 2014).
Increased both in the long- and the short-term.
C & E Europe, Black Sea & E Mediterranean (bre)
2000 - 2014
92,000 - 163,000
2000 - 2012
Increasing
CSR7
References
BirdLife International 2015. European Red List of Birds. Luxembourg: Office for Official Publications of the European Communities. http://datazone.birdlife.org/info/euroredlist
Notes
In WPE2 this population belonged to one single population (Europe (breeding)).
The population has increased in the short-term, but the short-term trend is unknown in 7 of 26 breeding range states. The long-term trend is possibly stable, but unknown in 10 of 26 range states.
South-west Asia (win)
2006 - 2006
25,000 - 100,000
2003 - 2012
Unknown
CSR7
References
Unpublished information supplied by Wetlands International Specialist Groups, 2006.
South Asia (non-bre)
—
—
—
Unknown
WPE5
S & E Asia (non-bre)
2001 - 2001
25,000 - 100,000
1981 - 1991
Declining
WPE5
References
Chan, Simba, In litt,. 2002. Unpublished information on Asian waterbirds.
Notes
Sometimes ascribed to orientalis.
capensis
Southern Africa
1980 - 2010
500 - 2,000
1980 - 2009
Declining
CSR7
References
Animal Demography Unit (2017) Southern African Bird Atlas Project 2. SABAP1 vs SABAP2 reporting rates (QDGC). URL: http://sabap2.adu.org.za/index.php
Dodman, T. 2014. Status, Estimates and Trends of Waterbird Populations in Africa: AEWA-listed African populations. Wetlands International. (CSR6 African populations) URL: https://www.wetlands.org/publications/1304/
Notes
Fragmented population, only low numbers assumed from any site.
Dodman (2014) assumed decline based on fragmented population and habitat loss in many areas. ADU (2017) data confirms that that the species was absent in 16 quarter degree grid cells in SABAP2 where it was present during SABAP1, declined in one and occupied only 5 new ones.
Garnett, S.T., Szabo, J.K. and Dutson, G. 2010. The Action Plan for Australian Birds 2010. CSIRO Publishing, Melbourne.
Pickering, R. and Gole, C. 2008. Swan coastal plain Australasian Bittern surveys 2007-2008. Report to Birds Australia, Perth.
Notes
This species was included as one population in the first edition and has now been divided into 4 smaller populations. (WPE2)
Estimate derived by WPE method of number of mature birds x1.5 and based on systematic range-wide surveys in recent years coordinated by BirdLife Australia.
Trend is declining (20-30% within 2 generations (11 years)) due to loss of habitat.
Garnett, S.T., Szabo, J.K. and Dutson, G. 2010. The Action Plan for Australian Birds 2010. CSIRO Publishing, Melbourne.
Notes
This species was included as one population in the first edition and has now been divided into 4 smaller populations. (WPE2)
Estimate derived by WPE method of number of mature birds x1.5 and based on systematic range-wide surveys in recent years coordinated by BirdLife Australia.
Trend is declining (20-30% within 2 generations (11 years)) due to loss of habitat.
New Zealand
2011 - 2011
1,000 - 1,500
2001 - 2010
Declining?
WPE5
References
O'Donnell, C. In litt. 30 March 2012 (advice from Department of Conservation, New Zealand).
Notes
This species was included as one population in the first edition and has now been divided into 4 smaller populations. (WPE2)
Estimate reflects better information arising from systematic investigation.
Trend assumed to be unchanged but requires confirmation.
New Caledonia
2011 - 2011
—
2001 - 2010
Extinct?
WPE5
References
Dutson, G. In litt. 21 March 2012b (advice regarding Pacific Islands including New Guinea).
Notes
This species was included as one population in the first edition and has now been divided into 4 smaller populations. (WPE2)
Botaurus lentiginosus
American Bittern
North America
2000 - 2000
2,980,000 - 2,980,000
1966 - 2004
Declining?
WPE5
References
Wiggins, D.A. (2006, September 6). American Bittern (Botaurus lentiginosus): a technical conservation assessment. [Online]. USDA Forest Service, Rocky Mountain Region.
Notes
Estimate from BBS 992,000 pairs (2,976,000 individuals). For populations over 2 million birds, Ramsar criterion 5 (20,000 or more waterbirds) applies.
Botaurus pinnatus
Pinnated Bittern
pinnatus
2005 - 2011
10,000 - 25,000
2001 - 2010
Stable
WPE5
References
Franke, I. 2006. Waterbirds in Per?: Final report. Museo de Historia Natural de la Universidad Nacional Mayor de San Marcos. Lima (unpublished report to BirdLife International).
Unterkofler, D. and Blanco, D.E. 2012. Unpublished information on Argentine waterbirds.
Wetlands International Argentina (compiler). 2012. Unpublished information from the Neotropical Waterbird Census, 1990-2011, including contributions from Colombia (Asociaci?n Calidris in litt.), Paraguay (Guyra Paraguay in litt.), Uruguay (Averaves in litt.) and Venezuela (Uni?n Venezolana de Ornit?logos in litt.).
Notes
No new data other than estimate of 576-2,900 for Central America (Komar et al. 2006).
caribaeus
—
—
—
Unknown
WPE5
Ixobrychus involucris
Stripe-backed Bittern
N South America
1993 - 1993
100,000 - 1,000,000
1982 - 1992
Stable
WPE5
References
Canevari, P. (pers. comm.). 1993. Unpublished information from Neotropical Wetlands Program.
Notes
In WPE2 this population belonged to one single population (South America).
S South America
2011 - 2011
10,000 - 25,000
2001 - 2010
Stable
WPE5
References
Huanca Llanos, N.E. 2006. Reporte final, Aves Acu?ticas en Bolivia. Asociaci?n Armonia, Santa Cruz de la Sierra (unpublished report to BirdLife International).
Unterkofler, D. and Blanco, D.E. 2012. Unpublished information on Argentine waterbirds.
Notes
In WPE2 this population belonged to one single population (South America).
Ixobrychus exilis
Least Bittern
exilis
2000 - 2000
128,000 - 128,000
—
Unknown
WPE5
References
Waterbird Conservation for the Americas. 2001. Continental Marshbird Conservation Planning Workshop, August 2001, Denver, Colorado.
Notes
No new data. 4,500 (1,500 Bpairs) in Canada (COSEWIC. 2009).
pullus
—
—
—
Unknown
WPE5
bogotensis
2011 - 2011
1,000 - 1,000
2001 - 2010
Declining
WPE5
References
Asociaci?n Calidris. 2012. Unpublished information from the Neotropical Waterbird Census in Colombia, 1992-2011.
erythromelas
2005 - 2011
10,000 - 25,000
—
Unknown
WPE5
References
Asociaci?n Calidris. 2012. Unpublished information from the Neotropical Waterbird Census in Colombia, 1992-2011.
peruvianus
2011 - 2011
1,000 - 5,000
2001 - 2010
Stable
WPE5
References
Ottema, O. 2006. Waterbirds in Suriname: Final report. STINASU Foundation for Nature Conservation Suriname, Paramaribo (unpublished report).
Unterkofler, D. and Blanco, D.E. 2012. Unpublished information on Argentine waterbirds.
Ixobrychus minutus
Common Little Bittern
minutus
W Europe, NW Africa/Subsaharan Africa
1997 - 2013
19,000 - 25,500
2000 - 2012
Stable
CSR7
References
BirdLife International 2015. European Red List of Birds. Luxembourg: Office for Official Publications of the European Communities. http://datazone.birdlife.org/info/euroredlist
Dodman, T. 2014. Status, Estimates and Trends of Waterbird Populations in Africa: AEWA-listed African populations. Wetlands International. (CSR6 African populations) URL: https://www.wetlands.org/publications/1304/
Notes
In WPE2 this population belonged to one single population (Europe/Northern Africa (bre)).
6,227-8253 pairs in BE, DE, ES, ESIC, FR, IT, LU, NL and PT (BirdLife International 2015). 100-200 pairs in NW Africa (Dodman, 2014).
Stable in the short-term but declined in the long-term.
C & E Europe, Black Sea & E Mediterranean/Sub-saharan Africa
1995 - 2014
168,000 - 298,000
2000 - 2012
Stable
CSR7
References
BirdLife International 2015. European Red List of Birds. Luxembourg: Office for Official Publications of the European Communities. http://datazone.birdlife.org/info/euroredlist
Dodman, T. 2014. Status, Estimates and Trends of Waterbird Populations in Africa: AEWA-listed African populations. Wetlands International. (CSR6 African populations) URL: https://www.wetlands.org/publications/1304/
15. eu
Notes
In WPE2 this population belonged to one single population (Europe/Northern Africa (bre)).
55,156 - 98,469 pairs in AL, AT, BA, BE, BG, BY, CH, CY, CZ, DE, ES, ESIC, FR, GE, GR, HR, HU, IT, LT, LU, LV, MD, ME, MK, NL, PL, PT, RO, RS, RU, SI, SK, TR, UA & XK (BirdLife International 2015). Further 1000 pairs in Egypt (Dodman, 2014).
Stable both in the long- and the short-term.
West & South-west Asia/Sub-Saharan Africa
1987 - 1991
25,000 - 100,000
2003 - 2012
Unknown
CSR7
References
Perennou, C.P., Mundkur, T. and Scott, D.A. 1994. The Asian Waterfowl Census 1987-1991: distribution and status of Asian waterfowl. IWRB Spec. Publ. No. 24; AWB Spec. Publ. No. 86. Slimbridge, UK and Kuala Lumpur, Malaysia.
South Asia
1987 - 1991
10,000 - 25,000
—
Unknown
WPE5
References
Perennou, C.P., Mundkur, T. and Scott, D.A. 1994. The Asian Waterfowl Census 1987-1991: distribution and status of Asian waterfowl. IWRB Spec. Publ. No. 24; AWB Spec. Publ. No. 86. Slimbridge, UK and Kuala Lumpur, Malaysia.
payesii
Sub-Saharan Africa
1990 - 2000
25,000 - 100,000
2003 - 2012
Unknown
CSR7
References
Fishpool, L.D.C. and Evans, M.I.,(Eds). 2001. Important Bird Areas in Africa and associated islands: Priority sites for conservation. Newbury and Cambridge, UK: Pisces Publications and BirdLife International (BirdLife Conservation Series No. 11).
podiceps
—
3,000 - 15,000
—
Unknown
WPE5
References
Hawkins, F. In litt. 2002. Unpublished notes on waterbird population estimates of Madagascar.
Ixobrychus dubius
Australian Little Bittern
dubius
1994 - 2010
1 - 10,000
1982 - 2011
Declining?
WPE5
References
Birds Australia. 2011. The Bittern Chronicle No. 3, September 2011. Online http://www.birdsaustralia.com.au/images/stories/current-projects/bittern/Bittern-Chronicle-Sep11.pdf accessed April 2012.
Christidis, L. and Boles, W.E. 2008. Systematics and taxonomy of Australian birds. CSIRO Publishing, Melbourne.
Clarke, A.G., Lane, J.A.K. and Jaensch, R.P. 2011. Surveys of waterbirds in selected wetlands of south-western Australia in spring-summer 2009-10, with further assessment of changes in habitat and waterbird usage over 2-3 decades. Western Australian Department of Environment and Conservation. 101pp.
Jaensch, R.P. 1995. Little Bitterns (Ixobrychus minutus) in the Middle Fly wetlands. Muruk 7 (3): 117-118.
Jaensch, Roger, August 2005. Detailed data and notes on numbers and distribution of waterbirds in Australia, updated from Jaensch, 2003, and derived from more than 90 Australian sources including Wetlands International, Birds Australia and Jaensch, 2003,
Jaensch, R. April 2012. Detailed data and notes on numbers and trends of waterbirds in Australia and other parts of the Oceania region, in a spreadsheet prepared for Wetlands International - Oceania for the 5th edition of WPE.
Notes
Now considered a distinct species, previously laced under Ixobrychus minutus
Data inadequate to derive a numerical estimate due to survey difficulties; numbers recorded support 1 - 10,000.
Trend is possibly declining due to loss of inland habitat.
Ixobrychus novaezelandiae
New Zealand Little Bittern
New Zealand
—
—
1982 - 1992
Extinct
WPE5
References
Powlesland, R. (pers. comm.). 1993. New Zealand Department of Conservation unpublished data.
Notes
Treated as subspecies of L.minutus until WPE4.
Ixobrychus sinensis
Yellow Bittern
Seychelles
—
1 - 300
1991 - 2001
Declining
WPE5
References
Rocamora, G. and Skerrett, A. 2001. Seychelles. Pp 751-768. In L.D.C. Fishpool and M.I. Evans, eds. Important Bird Areas in Africa and associated islands: Priority sites for conservation. Newbury and Cambridge, UK: Pisces Publications and BirdLife International
Skerrett, A., 2002. In litt.
South Asia
—
—
—
Unknown
WPE5
E & SE Asia
2006 - 2006
100,000 - 1,000,000
1985 - 2005
Increasing
WPE5
References
Unpublished information supplied by Wetlands International Specialist Groups, 2006.
Notes
42: Recently colonised New Caledonia.
Ixobrychus eurhythmus
Schrenck's Bittern
E & SE Asia
—
1 - 25,000
1985 - 1995
Declining
WPE5
References
Lansdown, R.V. (pers. comm.) 1996. Data prepared for the Draft Ardeidae Action Plan.
Ixobrychus cinnamomeus
Cinnamon Bittern
South Asia
1987 - 1991
25,000 - 1,000,000
—
Unknown
WPE5
References
Perennou, C.P., Mundkur, T. and Scott, D.A. 1994. The Asian Waterfowl Census 1987-1991: distribution and status of Asian waterfowl. IWRB Spec. Publ. No. 24; AWB Spec. Publ. No. 86. Slimbridge, UK and Kuala Lumpur, Malaysia.
E, SE Asia
2006 - 2006
100,000 - 1,000,000
1985 - 2005
Stable
WPE5
References
Unpublished information supplied by Wetlands International Specialist Groups, 2006.
Ixobrychus sturmii
Dwarf Bittern
Sub-Saharan Africa
1990 - 2000
25,000 - 100,000
2002 - 2013
Unknown
CSR7
References
Fishpool, L.D.C. and Evans, M.I.,(Eds). 2001. Important Bird Areas in Africa and associated islands: Priority sites for conservation. Newbury and Cambridge, UK: Pisces Publications and BirdLife International (BirdLife Conservation Series No. 11).
Ixobrychus flavicollis
Black Bittern
flavicollis
South Asia
1987 - 1991
25,000 - 100,000
—
Unknown
WPE5
References
Perennou, C.P., Mundkur, T. and Scott, D.A. 1994. The Asian Waterfowl Census 1987-1991: distribution and status of Asian waterfowl. IWRB Spec. Publ. No. 24; AWB Spec. Publ. No. 86. Slimbridge, UK and Kuala Lumpur, Malaysia.
Notes
Sometimes ascribed to genus Dupetor.
E, SE Asia
2006 - 2006
10,000 - 100,000
—
Unknown
WPE5
References
Unpublished information supplied by Wetlands International Specialist Groups, 2006.
australis
Australia, New Guinea
1999 - 2002
10,000 - 100,000
1991 - 2000
Declining
WPE5
References
Bishop, K.D. 2003. A Review of the Avifauna of the TransFly Eco-region: the status, distribution, habitats and conservation of the region?s birds. Report to WWF South Pacific Program.
Green, A.J. 1992a. The status and conservation of the White-winged Wood Duck (Cairina scutulata). IWRB Special Publication No. 17. Slimbridge, UK. 115 pp.
Jaensch, R. April 2012. Detailed data and notes on numbers and trends of waterbirds in Australia and other parts of the Oceania region, in a spreadsheet prepared for Wetlands International - Oceania for the 5th edition of WPE.
Notes
Sometimes ascribed to gouldi.
Estimate based on previous information and extent of habitat across several countries.
Timor
—
—
—
Unknown
WPE5
Rennell Island
2001 - 2001
1 - 500
—
Unknown
WPE5
References
Dutson, Guy, in litt. 2002.
Notes
Sometimes included in woodfordi, or as separate subspecies, pallidor.
New Britain, New Ireland
2001 - 2001
1 - 10,000
—
Unknown
WPE5
References
Dutson, Guy, in litt. 2002.
Notes
Sometimes ascribed to nesophilus.
woodfordi
2001 - 2001
1 - 10,000
—
Unknown
WPE5
References
Dutson, Guy, in litt. 2002.
Gorsachius magnificus
White-eared Night-heron
SE Asia
2000 - 2000
250 - 1,000
1995 - 2005
Declining
WPE5
References
BirdLife International 2001. Threatened Bird of Asia: the BirdLife International Red Data Book. Cambridge, UK. BirdLife International.
BirdLife International. 2005. Threatened birds of the world 2005. Species factsheets available at?www.birdlife.org
Notes
M. Crosby in litt. (2005) estimates 250-999 individuals based on analysis of information presented in 66. Habitat detrimentally transformed at principal site, Shennongjia in Hubei province, China, in 1990s.
Gorsachius goisagi
Japanese Night-heron
E & SE Asia
2005 - 2005
250 - 1,000
1975 - 1995
Declining
WPE5
References
BirdLife International. 2005. Threatened birds of the world 2005. Species factsheets available at?www.birdlife.org
Gorsachius melanolophus
Malay Night-heron
South Asia
—
—
—
Unknown
WPE5
SE Asia
—
—
—
Unknown
WPE5
Philippines
—
—
—
Unknown
WPE5
Notes
Sometimes ascribed to kutteri. Probably this subspecies on Ryukyu Is and Taiwan.
Palawan, Philippines
—
1 - 10,000
—
Unknown
WPE5
References
Lansdown, R.V. (pers. comm.) 1996. Data prepared for the Draft Ardeidae Action Plan.
Notes
Sometimes ascribed to rufolineatus.
Nicobar Is
—
1 - 10,000
—
Unknown
WPE5
References
Lansdown, R.V. (pers. comm.) 1996. Data prepared for the Draft Ardeidae Action Plan.
Notes
Sometimes ascribed to minor.
Calherodius leuconotus
White-backed Night-heron
Africa
—
25,000 - 100,000
1991 - 2001
Stable
WPE5
References
Dodman, T. 2002. Waterbird Population Estimates in Africa. Unpublished report to Wetlands International.
Dodman, T., 2006. Status, estimates and trends of waterbird populations in Africa. Wetlands International, Dakar.
Notes
Sometimes included in genus Gorsachius.
Nycticorax duboisi
Reunion Night-heron
Reunion
2005 - 2005
—
1750 - 2005
Extinct
WPE5
References
BirdLife International. 2005. Threatened birds of the world 2005. Species factsheets available at?www.birdlife.org
Notes
Population added in WPE3.
Described and last recorded in 1674 (BirdLife International. 2005).
Nycticorax mauritianus
Mauritius Night-heron
Mauritius
2005 - 2005
—
1750 - 2005
Extinct
WPE5
References
BirdLife International. 2005. Threatened birds of the world 2005. Species factsheets available at?www.birdlife.org
Notes
Population added in WPE3.
Described and last recorded in 1693 (BirdLife International. 2005).
Nycticorax megacephalus
Rodrigues Night-heron
Rodrigues
2005 - 2005
—
1800 - 2005
Extinct
WPE5
References
BirdLife International. 2005. Threatened birds of the world 2005. Species factsheets available at?www.birdlife.org
Notes
Population added in WPE3.
Described and last recorded in 1726 (BirdLife International. 2005).
Nycticorax nycticorax
Black-crowned Night-heron
nycticorax
W Europe, NW Africa (bre)
2002 - 2012
46,000 - 51,000
1998 - 2012
Declining
CSR7
References
BirdLife International 2015. European Red List of Birds. Luxembourg: Office for Official Publications of the European Communities. http://datazone.birdlife.org/info/euroredlist
Dodman, T. 2014. Status, Estimates and Trends of Waterbird Populations in Africa: AEWA-listed African populations. Wetlands International. (CSR6 African populations) URL: https://www.wetlands.org/publications/1304/
Notes
In WPE2 this population belonged to one single population (Europe/NW Africa (breeding)).
14,836 - 15,596 pairs in BE, DE, ES, ESIC , FR, IT, NL and PT (BirdLife International 2015). Dodman (2014) estimated that 500-1500 pairs may breed in NW Africa.
The European part of the population has declined by 50-53% over the last decade.
C & E Europe/Black Sea & E Mediterranean (bre)
1990 - 2012
134,000 - 209,000
2000 - 2012
Stable
CSR7
References
BirdLife International 2015. European Red List of Birds. Luxembourg: Office for Official Publications of the European Communities. http://datazone.birdlife.org/info/euroredlist
Dodman, T. 2014. Status, Estimates and Trends of Waterbird Populations in Africa: AEWA-listed African populations. Wetlands International. (CSR6 African populations) URL: https://www.wetlands.org/publications/1304/
Notes
In WPE2 this population belonged to one single population (Europe/NW Africa (breeding)).
44,700 - 69,610 pairs in AL, AT, AZ, BA, BG, BY, GE, GR, HR, HU, MD, ME, MK, PL, RO, RS, RU, SI, SK, TR, UA& XK (BirdLife International 2015). In addition, less than 1000 birds in Egypt (Dodman, 2014).
Stable both in the short- and the long-term.
Western Asia/SW Asia & NE Africa
2002 - 2017
25,000 - 100,000
2000 - 2012
Unknown
CSR7
References
Scott, D.A. in press, 2002. Report on the Conservation Status of Migratory Waterbirds in the Agreement Area. Update Report to African Eurasian Migratory Waterbird Agreement Secretariat
Sheldon, R. 2017. Estimates of breeding waterbird populations in Central/SW Asia, The Caucasus and the Arabian Peninsula.
Sub-Saharan Africa & Madagascar
1975 - 2014
100,000 - 300,000
2006 - 2015
Stable/ Increasing?
CSR7
References
Dodman, T. 2014. Status, Estimates and Trends of Waterbird Populations in Africa: AEWA-listed African populations. Wetlands International. (CSR6 African populations) URL: https://www.wetlands.org/publications/1304/
Wetlands International (2017) Flyway trend analyses based on data from the African-Eurasian Waterbird Census from the period of 1967-2015. Ede, The Netherlands: Wetlands International. URL: http://iwc.wetlands.org/index.php/aewatrends
Notes
Widespread, with breeding colonies across sub-Saharan Africa
Dodman (2014) considered it to be at least stable. IWC trend analysis shows strong increase both in the long- and short-term, but this is driven by data from SN (Wetlands International 2017)
South Asia
—
100,000 - 150,000
1977 - 1991
Stable
WPE5
References
Balachandran, S., in litt. 2005.
Perennou, C.P., Mundkur, T. and Scott, D.A. 1994. The Asian Waterfowl Census 1987-1991: distribution and status of Asian waterfowl. IWRB Spec. Publ. No. 24; AWB Spec. Publ. No. 86. Slimbridge, UK and Kuala Lumpur, Malaysia.
Notes
33: Population estimate for India 100,000.
E, SE Asia
2006 - 2006
100,000 - 1,000,000
1981 - 1991
Stable
WPE5
References
Chan, Simba, In litt,. 2002. Unpublished information on Asian waterbirds.
Unpublished information supplied by Wetlands International Specialist Groups, 2006.
In WPE2 this population belonged to one single population (N.n. nycticorax, North America/N Neotropics).
Total of data presented in BirdLife International 2008a.
northern South America (bre)
2005 - 2010
50,000 - 50,000
2001 - 2010
Stable
WPE5
References
Huanca Llanos, N.E. 2006. Reporte final, Aves Acu?ticas en Bolivia. Asociaci?n Armonia, Santa Cruz de la Sierra (unpublished report to BirdLife International).
Unterkofler, D. and Blanco, D.E. 2012. Unpublished information on Argentine waterbirds.
Wetlands International Argentina (compiler). 2012. Unpublished information from the Neotropical Waterbird Census, 1990-2011, including contributions from Colombia (Asociaci?n Calidris in litt.), Paraguay (Guyra Paraguay in litt.), Uruguay (Averaves in litt.) and Venezuela (Uni?n Venezolana de Ornit?logos in litt.).
Notes
In WPE2 this population belonged to one single population (N.n. nycticorax, North America/N Neotropics).
North America (bre)
1999 - 2011
75,000 - 150,000
1990 - 2000
Stable
WPE5
References
Butler, R.W., Kushlan, J.A. and Davidson, I.J. 2000. Herons in North America, Central America and the West Indies. Pp 151-175 in: Heron Conservation (James.A. Kushlan and Heinz Hafner, Eds.). Academic Press, London.
Schlatter, R. (In litt.). 2002. Unpublished information on Southern South American waterbirds.
Notes
Ascribed to subspecies hoactli (J. F. Gmelin, 1789) . In WPE2 this population belonged to one single population (N.n. nycticorax, North America/N Neotropics).
2,500-5,000 breeding pairs estimated for Canada (Weseloh, C. In litt. 2012).
Ascribed to subspecies hoactli (J. F. Gmelin, 1789).
Mexico
—
—
—
Unknown
WPE5
Notes
In WPE2 this population belonged to one single population (N.n. nycticorax, North America/N Neotropics).
Central America
2005 - 2005
9,600 - 24,900
—
Unknown
WPE5
References
Komar, O., Angehr, G., Eisermann, K., Herrera, N. and Zolotoff, J.M.. 2006. Waterbird Population Estimates for Central America. IV North American Ornithological Conference. unpublished report.
Notes
In WPE2 this population belonged to one single population (N.n. nycticorax, North America/N Neotropics).
obscurus
2001 - 2001
25,000 - 1,000,000
—
Unknown
WPE5
References
Schlatter, R. (In litt.). 2002. Unpublished information on Southern South American waterbirds.
falklandicus
Falkland Islands (Malvinas)
—
5,700 - 10,800
—
Unknown
WPE5
References
Woods, R. and Woods, A. 1997. Atlas of the Breeding Birds of the Falkland Islands. Anthony Nelson, Oswestry, England.
Barr?, N. and Dutson, G. 2000. Oiseaux de Nouvelle-Caledonie. Liste commentee. Suppl. Alauda (68), 3: 49pp.
manillensis
—
—
—
Unknown
WPE5
Notes
Includes former N.caledonicus minahassae.
pelewensis
2001 - 2001
1 - 10,000
—
Unknown
WPE5
References
Dutson, Guy, in litt. 2002.
mandibularis
2001 - 2001
10,000 - 100,000
—
Unknown
WPE5
References
Dutson, Guy, in litt. 2002.
Notes
Bismarck birds sometimes separated as cancrivorus.
hilli
2004 - 2011
25,000 - 1,000,000
1982 - 2011
Fluctuating
WPE5
References
Jaensch, Roger, August 2005. Detailed data and notes on numbers and distribution of waterbirds in Australia, updated from Jaensch, 2003, and derived from more than 90 Australian sources including Wetlands International, Birds Australia and Jaensch, 2003,
Jaensch, R. April 2012. Detailed data and notes on numbers and trends of waterbirds in Australia and other parts of the Oceania region, in a spreadsheet prepared for Wetlands International - Oceania for the 5th edition of WPE.
Reid, J. and Jaensch, R. 2004. Aerial waterbird survey results. Section 4.7, pp 472-493 in, Costelloe, J.F., Hudson, P.J., Pritchard, J.C., Puckridge, J.T. and Reid, J.R.W. AridFlo Scientific Report: Environmental Flow Requirements of Arid Zone Rivers with Particular Reference to the Lake Eyre Drainage Basin. School of Earth and Environmental Sciences, University of Adelaide, Adelaide. Final Report to DLW and BC and DEH. Also available online at http://www.lebmf.gov.au/publications/aridflo.html
Reid, J.R.W., Kingsford, R.T. and Jaensch, R.P. 2009. Waterbird Surveys in the Channel Country Floodplain Wetlands, Autumn 2009. Report by Australian National University, Canberra, University of New South Wales, Sydney, and Wetlands International, Oceania, Brisbane, for the Australian Government Department of Environment, Water, Heritage and the Arts. 81 pp. Also available online at http://www.lebmf.gov.au/publications/pubs/waterbirds-report.pdf
Notes
Estimate based on numbers in colonies and consideration of survey difficulties and numbers outside Australia.
Trend is fluctuating; possible long-term decline due to loss of some breeding habitat in E Australia requires confirmation.
Nyctanassa violacea
Yellow-crowned Night-heron
violacea
North America
—
75,000 - 150,000
1966 - 2000
Stable
WPE5
References
Butler, R.W., Kushlan, J.A. and Davidson, I.J. 2000. Herons in North America, Central America and the West Indies. Pp 151-175 in: Heron Conservation (James.A. Kushlan and Heinz Hafner, Eds.). Academic Press, London.
Sauer, J. R., J. E. Hines, and J. Fallon. 2001. The North American Breeding Bird Survey, Results and Analysis 1966 - 2000. Version 2001.2, USGS Patuxent Wildlife Research Center, Laurel, MD.
Notes
Often placed in genus Nycticorax. In WPE2 this population belonged to one single population (N.v. violacea, North Am/Central America).
Mexico to Honduras
—
—
—
Unknown
WPE5
Notes
In WPE2 this population belonged to one single population (N.v. violacea, North Am/Central America).
No new data other than estimate of 1,400-1,900 for Central America (Komar et al. 2006).
gravirostris
—
—
—
Unknown
WPE5
Notes
Sometimes ascribed to bancrofti.
caliginis
2004 - 2004
900 - 900
1982 - 1992
Stable
WPE5
References
Angehr, G. R. 2005. Waterbirds in Panama. Final report to BirdLife International.
Naranjo, L. (pers. comm.). 1993. Unpublished data summarised for IWRB.
cayennensis
—
—
—
Unknown
WPE5
bancrofti
—
—
—
Unknown
WPE5
Notes
No new data other than estimate of 2,203-7,603 for Caribbean (Birdlife International 2008a).
pauper
—
—
—
Unknown
WPE5
Butorides striata
Green-backed Heron
brevipes
—
1 - 25,000
—
Unknown
CSR6
References
Jennings, M.C. 2010. Atlas of Breeding Birds of Arabia. Fauna of Arabia Vol. 25. Senckenberg Gesellschaft für Naturforschung and King Abdulaziz City for Science and Technology, Frankfurt, Germany and Riyadh, Saudi Arabia.
Notes
Jennings et al. estimated that the population is 2000 pairs in Arabia.
sundevalli
—
—
—
Unknown
WPE5
Notes
Sometimes considered separate species Butorides sundevalli.
albolimbata
—
1 - 10,000
—
Unknown
WPE5
References
Lansdown, R.V. (pers. comm.) 1996. Data prepared for the Draft Ardeidae Action Plan.
chloriceps
—
25,000 - 25,000
1991 - 2001
Declining
WPE5
References
Hoffman, T.W., Deepal Warakagoda, U. Sirivardena, 2002. Unpublished information on the birds of Sri Lanka.
spodiogaster
—
—
—
Unknown
WPE5
amurensis
—
—
—
Unknown
WPE5
actophila
—
—
—
Unknown
WPE5
Note
Includes "connectens". (WPE4)
javanica
2006 - 2006
10,000 - 100,000
—
Unknown
WPE5
References
Unpublished information supplied by Wetlands International Specialist Groups, 2006.
Note
Split into a SE & S Asia population and a Mauritius, Reunion & Rodrigues population after WPE5.
moluccarum
Moluccas
—
—
—
Unknown
WPE5
NW New Guinea
—
—
—
Unknown
WPE5
Notes
Sometimes ascribed to papuensis.
solomonensis
2001 - 2001
1 - 25,000
—
Unknown
WPE5
References
Dutson, Guy, in litt. 2002.
atricapilla
2001 - 2001
100,000 - 1,000,000
1991 - 2001
Stable
WPE5
References
Dodman, T. 2002. Waterbird Population Estimates in Africa. Unpublished report to Wetlands International.
idenburgi
—
—
—
Unknown
WPE5
flyensis
—
—
—
Unknown
WPE5
stagnatilis
N Australia
—
—
—
Unknown
WPE5
NW Australia
—
—
—
Unknown
WPE5
Notes
Sometimes ascribed to rogersi.
NE Western Australia
—
—
—
Unknown
WPE5
Notes
Sometimes ascribed to cinereus.
patruelis
—
100 - 200
1971 - 1987
Declining
WPE5
References
Thibault, J.C. and Guyst, I. 1993. Livre rouge des oiseaux menac?s des regions Francaises d'outre mer. ICBP Monograph No. 5. CIPO, Saint-Claude, France.
macrorhyncha
E Australia
—
—
—
Unknown
WPE5
SC New Guinea, NE Queensland
—
—
—
Unknown
WPE5
Notes
Sometimes ascribed to littleri.
carcinophila
—
—
—
Unknown
WPE5
steini
—
—
—
Unknown
WPE5
rutenbergi
2001 - 2001
1 - 25,000
1991 - 2001
Stable
WPE5
References
Dodman, T. 2002. Waterbird Population Estimates in Africa. Unpublished report to Wetlands International.
rhizophorae
2001 - 2001
1 - 10,000
1992 - 2002
Stable
WPE5
References
Dodman, T. 2002. Waterbird Population Estimates in Africa. Unpublished report to Wetlands International.
Louette, M. 2004. Oiseaux. In: Louette, M., D. Meirte and R. Jocque (eds). La faune terrestre de l?archipel des Comores. Studies in Afrotropical Zoology (MRAC, Tervuren). 293: 89-196.
degens
2001 - 2001
1 - 10,000
—
Unknown
WPE5
References
Dodman, T. 2002. Waterbird Population Estimates in Africa. Unpublished report to Wetlands International.
crawfordi
2001 - 2001
1 - 5,000
1991 - 2001
Stable
WPE5
References
Dodman, T. 2002. Waterbird Population Estimates in Africa. Unpublished report to Wetlands International.
striata
Central & South America
—
—
—
Unknown
WPE5
South America
2005 - 2011
100,000 - 1,000,000
2001 - 2010
Stable
WPE5
References
Huanca Llanos, N.E. 2006. Reporte final, Aves Acu?ticas en Bolivia. Asociaci?n Armonia, Santa Cruz de la Sierra (unpublished report to BirdLife International).
Ottema, O. 2006. Waterbirds in Suriname: Final report. STINASU Foundation for Nature Conservation Suriname, Paramaribo (unpublished report).
Unterkofler, D. and Blanco, D.E. 2012. Unpublished information on Argentine waterbirds.
Wetlands International Argentina (compiler). 2012. Unpublished information from the Neotropical Waterbird Census, 1990-2011, including contributions from Colombia (Asociaci?n Calidris in litt.), Paraguay (Guyra Paraguay in litt.), Uruguay (Averaves in litt.) and Venezuela (Uni?n Venezolana de Ornit?logos in litt.).
Notes
Sometimes ascribed to cyanurus.
virescens
Central & E North America
—
—
1966 - 2000
Declining
WPE5
References
Sauer, J. R., J. E. Hines, and J. Fallon. 2001. The North American Breeding Bird Survey, Results and Analysis 1966 - 2000. Version 2001.2, USGS Patuxent Wildlife Research Center, Laurel, MD.
Notes
Butorides striata (del Hoyo and Collar 2014) was previously split as B. striata and B. virescens
Mexico
—
—
—
Unknown
WPE5
Notes
Butorides striata (del Hoyo and Collar 2014) was previously split as B. striata and B. virescens. Sometimes ascribed to maculatus. (WPE4)
Central America
2005 - 2005
23,000 - 60,000
—
Unknown
WPE5
References
Komar, O., Angehr, G., Eisermann, K., Herrera, N. and Zolotoff, J.M.. 2006. Waterbird Population Estimates for Central America. IV North American Ornithological Conference. unpublished report.
Notes
Butorides striata (del Hoyo and Collar 2014) was previously split as B. striata and B. virescens
Butorides striata (del Hoyo and Collar 2014) was previously split as B. striata and B. virescens
Total of data presented in BirdLife International 2008a
bahamensis
—
—
—
Unknown
WPE5
Notes
Butorides striata (del Hoyo and Collar 2014) was previously split as B. striata and B. virescens, though this population was considered Butorides striatus in WPE1.
anthonyi
—
—
1966 - 2000
Increasing
WPE5
References
Sauer, J. R., J. E. Hines, and J. Fallon. 2001. The North American Breeding Bird Survey, Results and Analysis 1966 - 2000. Version 2001.2, USGS Patuxent Wildlife Research Center, Laurel, MD.
Notes
Butorides striata (del Hoyo and Collar 2014) was previously split as B. striata and B. virescens. This population was under Butorides striatus in WPE1.
frazari
—
—
—
Unknown
WPE5
Notes
Butorides striata (del Hoyo and Collar 2014) was previously split as B. striata and B. virescens. This population was considered Butorides striatus in WPE1.
Ardeola ralloides
Squacco Heron
ralloides
SW Europe, NW Africa (bre)
2002 - 2013
9,000 - 11,000
2000 - 2012
Increasing
CSR7
References
BirdLife International 2015. European Red List of Birds. Luxembourg: Office for Official Publications of the European Communities. http://datazone.birdlife.org/info/euroredlist
Dodman, T. 2014. Status, Estimates and Trends of Waterbird Populations in Africa: AEWA-listed African populations. Wetlands International. (CSR6 African populations) URL: https://www.wetlands.org/publications/1304/
Notes
8,495-10,703 pairs in ES, FR, IT & PT (BirdLife International 2015). C. 100 pairs in N. Africa (Dodman, 2014).
Increased both in the short- and the long-term.
C & E Europe, Black Sea & E Mediterranean (bre)
1990 - 2012
29,000 - 52,000
2000 - 2012
Declining
CSR7
References
BirdLife International 2015. European Red List of Birds. Luxembourg: Office for Official Publications of the European Communities. http://datazone.birdlife.org/info/euroredlist
Dodman, T. 2014. Status, Estimates and Trends of Waterbird Populations in Africa: AEWA-listed African populations. Wetlands International. (CSR6 African populations) URL: https://www.wetlands.org/publications/1304/
Notes
In WPE2 this population belonged to one single population (S&SW Asia/Black Sea (bre)).
9,219-16,569 pairs in AL, BA, BG, CY, GE, GR, HR, HU, MD, ME, MK, RO, RS, RU, SK, TR & UA (BirdLife International 2015). In addition, over 600 breeding pairs in Egypt (Dodman, 2014).
Declining both in the long- and the short-term.
West & South-west Asia/Sub-Saharan Africa
1987 - 2017
25,000 - 100,000
2003 - 2012
Unknown
CSR7
References
BirdLife International Website 2002
Perennou, C.P., Mundkur, T. and Scott, D.A. 1994. The Asian Waterfowl Census 1987-1991: distribution and status of Asian waterfowl. IWRB Spec. Publ. No. 24; AWB Spec. Publ. No. 86. Slimbridge, UK and Kuala Lumpur, Malaysia.
Sheldon, R. 2017. Estimates of breeding waterbird populations in Central/SW Asia, The Caucasus and the Arabian Peninsula.
Notes
In WPE2 this population belonged to one single population (S&SW Asia/Black Sea (bre)).
paludivaga
Sub-Saharan Africa & Madagascar
2006 - 2006
300,000 - 600,000
2006 - 2015
Increasing?
CSR7
References
Dodman, T., 2006. Status, estimates and trends of waterbird populations in Africa. Wetlands International, Dakar.
Trolliet, B., in litt., 2006.
Wetlands International (2017) Flyway trend analyses based on data from the African-Eurasian Waterbird Census from the period of 1967-2015. Ede, The Netherlands: Wetlands International. URL: http://iwc.wetlands.org/index.php/aewatrends
Notes
Sometimes ascribed to ralloides.
Ardeola grayii
Indian Pond-heron
grayii
SW, S Asia
1987 - 1991
100,000 - 1,000,000
—
Unknown
WPE5
References
Perennou, C.P., Mundkur, T. and Scott, D.A. 1994. The Asian Waterfowl Census 1987-1991: distribution and status of Asian waterfowl. IWRB Spec. Publ. No. 24; AWB Spec. Publ. No. 86. Slimbridge, UK and Kuala Lumpur, Malaysia.
Myanmar, Bay of Bengal
—
—
—
Unknown
WPE5
phillipsi
—
—
—
Unknown
WPE5
Ardeola bacchus
Chinese Pond-heron
E, SE & S Asia
2001 - 2001
25,000 - 1,000,000
1981 - 1991
Stable
WPE5
References
Chan, Simba, In litt,. 2002. Unpublished information on Asian waterbirds.
Ardeola speciosa
Javan Pond-heron
speciosa
—
—
—
Unknown
WPE5
continentalis
1987 - 1991
10,000 - 100,000
—
Unknown
WPE5
References
Perennou, C.P., Mundkur, T. and Scott, D.A. 1994. The Asian Waterfowl Census 1987-1991: distribution and status of Asian waterfowl. IWRB Spec. Publ. No. 24; AWB Spec. Publ. No. 86. Slimbridge, UK and Kuala Lumpur, Malaysia.
Ardeola idae
Madagascar Pond-heron
Madagascar & Aldabra/Central & Eastern Africa
2001 - 2001
2,000 - 6,000
2002 - 2013
Increasing?
CSR7
References
BirdLife International (2017) IUCN Red List for birds. Downloaded from http://www.birdlife.org on 30/09/2017.
Dodman, T. 2002. Waterbird Population Estimates in Africa. Unpublished report to Wetlands International.
Wetlands International (2017) Flyway trend analyses based on data from the African-Eurasian Waterbird Census from the period of 1967-2015. Ede, The Netherlands: Wetlands International. URL: http://iwc.wetlands.org/index.php/aewatrends
Notes
Significant long-term decline. BirdLife International (2017) suspects that the decline continues. However, IWC count data suggest modest increase after 2000 (Wetlands International 2017).
Ardeola rufiventris
Rufous-bellied Heron
Central, Eastern & Southern Africa
2006 - 2006
10,000 - 100,000
2006 - 2015
Stable/ Increasing?
CSR7
References
Unpublished information supplied by Wetlands International Specialist Groups, 2006
Wetlands International (2017) Flyway trend analyses based on data from the African-Eurasian Waterbird Census from the period of 1967-2015. Ede, The Netherlands: Wetlands International. URL: http://iwc.wetlands.org/index.php/aewatrends
Bubulcus ibis
Cattle Egret
ibis
Southern Africa
1996 - 2001
100,000 - 1,000,000
2006 - 2015
Declining?
CSR7
References
Animal Demography Unit (2017) Southern African Bird Atlas Project 2. SABAP1 vs SABAP2 reporting rates (QDGC). URL: http://sabap2.adu.org.za/index.php
Scott, D.A. in press, 2002. Report on the Conservation Status of Migratory Waterbirds in the Agreement Area. Update Report to African Eurasian Migratory Waterbird Agreement Secretariat.
Wetlands International (2017) Flyway trend analyses based on data from the African-Eurasian Waterbird Census from the period of 1967-2015. Ede, The Netherlands: Wetlands International. URL: http://iwc.wetlands.org/index.php/aewatrends
Notes
Often placed in genus Ardea.
The short-term trend has a strong negative tendency. The long-term trend is also significant long-term decline (Wetlands International 2017). It has declined in three times more quarter degree grid cells than increased in ZA between the SABAP 1 and 2 (ADU 2017).
In WPE2 this population belonged to one single population (B.i. ibis, Neotropics).Often placed in genus Ardea.
Total of data presented in BirdLife International 2008a and of which 25,000-100,000 estimated for Cuba (Acosta-Cruz, M. and Mugica-Vald?s, L. 2006.)
South America
1993 - 1993
1,000,000 - 1,000,000
1982 - 1992
Increasing
WPE5
References
Canevari, P. (pers. comm.). 1993. Unpublished information from Neotropical Wetlands Program.
Naranjo, L. (pers. comm.). 1993. Unpublished data summarised for IWRB.
Notes
In WPE2 this population belonged to one single population (B.i. ibis, Neotropics).Often placed in genus Ardea.
Tropical Africa
1990 - 2001
1,000,000 - 10,000,000
2006 - 2015
Unknown
CSR7
References
Dodman, T. 2014. Status, Estimates and Trends of Waterbird Populations in Africa: AEWA-listed African populations. Wetlands International. (CSR6 African populations) URL: https://www.wetlands.org/publications/1304/
Scott, D.A. in press, 2002. Report on the Conservation Status of Migratory Waterbirds in the Agreement Area. Update Report to African Eurasian Migratory Waterbird Agreement Secretariat.
Wetlands International (2017) Flyway trend analyses based on data from the African-Eurasian Waterbird Census from the period of 1967-2015. Ede, The Netherlands: Wetlands International. URL: http://iwc.wetlands.org/index.php/aewatrends
Notes
Often placed in genus Ardea.
Population probably numbers 'several million' (Dodman, 2014).
IWC trend analysis produced uncertain results.
North-west Africa
1984 - 2000
100,000 - 150,000
2006 - 2015
Stable?
CSR7
References
Hafner, H. 2000. Herons in The Mediterranean. Pp 32-54 in: Heron Conservation (James.A. Kushlan and Heinz Hafner, Eds.). Academic Press, London.
Wetlands International (2017) Flyway trend analyses based on data from the African-Eurasian Waterbird Census from the period of 1967-2015. Ede, The Netherlands: Wetlands International. URL: http://iwc.wetlands.org/index.php/aewatrends
Notes
In WPE2 this population belonged to one single population (SW Europe/NW Africa).Often placed in genus Ardea.
Stable long-term trend.
South-west Europe
2002 - 2012
215,000 - 253,000
2000 - 2012
Declining?
CSR7
References
BirdLife International 2015. European Red List of Birds. Luxembourg: Office for Official Publications of the European Communities. http://datazone.birdlife.org/info/euroredlist
Wetlands International (2017) Flyway trend analyses based on data from the African-Eurasian Waterbird Census from the period of 1967-2015. Ede, The Netherlands: Wetlands International. URL: http://iwc.wetlands.org/index.php/aewatrends
Notes
In WPE2 this population belonged to one single population (SW Europe/NW Africa).Often placed in genus Ardea.
71,770 - 84,193 pairs.
Both the breeding (BirdLife International 2015) and the IWC data (Wetlands International 2017) indicate long-term increase that turned into a decline in the short-term.
East Mediterranean & South-west Asia
2005 - 2017
10,000 - 100,000
2006 - 2015
Unknown
CSR7
References
Sheldon, R. 2017. Estimates of breeding waterbird populations in Central/SW Asia, The Caucasus and the Arabian Peninsula.
Wetlands International (2017) Flyway trend analyses based on data from the African-Eurasian Waterbird Census from the period of 1967-2015. Ede, The Netherlands: Wetlands International. URL: http://iwc.wetlands.org/index.php/aewatrends
Notes
Often placed in genus Ardea.
Although the trend analysis suggest steep decline, Hatzofe (pers. com) indicated that the species has exploded in IL.
North America
—
1,000,000 - 1,000,000
1966 - 2000
Stable
WPE5
References
Kushlan, James, A., Melanie J. Steinkamp, Katharine Parsons, Jack Capp, Martin Acosta Cruz, Malcolm Coulter, Ian Davidson, Loney Dickson, Naomi Edelson, Richard Elliot, R. Michael Erwin, Scott Hatch, Stephen Kress, Robert Milko, Steve Miller, Kyra Mills, Richard Paul, Roberto Phillips, Jorge E. Saliva, Bill Sydeman, John Trapp, Jennifer Wheeler, and Kent Wohl. 2002. North American Waterbird Conservation Plan, Version 1. Waterbird Conservation for the Americas, Washington, DC, USA.
Sauer, J. R., J. E. Hines, and J. Fallon. 2001. The North American Breeding Bird Survey, Results and Analysis 1966 - 2000. Version 2001.2, USGS Patuxent Wildlife Research Center, Laurel, MD.
Notes
Often placed in genus Ardea.
Apparently does not breed in Canada anymore, certainly not in Ontario (Weseloh, C. In litt. 2012).
Mexico
—
—
—
Unknown
WPE5
Notes
In WPE2 this population belonged to one single population (B.i. ibis, Neotropics).Often placed in genus Ardea.
Central America
2005 - 2005
262,000 - 2,120,000
2005 - 2005
Increasing
WPE5
References
Komar, O., Angehr, G., Eisermann, K., Herrera, N. and Zolotoff, J.M.. 2006. Waterbird Population Estimates for Central America. IV North American Ornithological Conference. unpublished report.
Notes
In WPE2 this population belonged to one single population (B.i. ibis, Neotropics).Often placed in genus Ardea.
coromanda
South Asia
1987 - 1991
100,000 - 1,000,000
—
Unknown
WPE5
References
Perennou, C.P., Mundkur, T. and Scott, D.A. 1994. The Asian Waterfowl Census 1987-1991: distribution and status of Asian waterfowl. IWRB Spec. Publ. No. 24; AWB Spec. Publ. No. 86. Slimbridge, UK and Kuala Lumpur, Malaysia.
Notes
Often placed in genus Ardea.
E, SE Asia
2001 - 2001
100,000 - 1,000,000
1981 - 1991
Stable
WPE5
References
Chan, Simba, In litt,. 2002. Unpublished information on Asian waterbirds.
Notes
Often placed in genus Ardea.
Oceania
1999 - 2011
25,000 - 1,000,000
1982 - 2011
Increasing
WPE5
References
Chatto, R. 2000. Waterbird breeding colonies in the Top End of the Northern Territory. Technical Report 69, Parks and Wildlife Commission of the Northern Territory, Australia.
Jaensch, Roger, August 2005. Detailed data and notes on numbers and distribution of waterbirds in Australia, updated from Jaensch, 2003, and derived from more than 90 Australian sources including Wetlands International, Birds Australia and Jaensch, 2003,
Jaensch, R. April 2012. Detailed data and notes on numbers and trends of waterbirds in Australia and other parts of the Oceania region, in a spreadsheet prepared for Wetlands International - Oceania for the 5th edition of WPE.
Notes
Madsen, J., Reed, A. and Andreev, A. 1996. Status and trends of geese (Anser sp., Branta sp.) in the world: a review, updating and evaluation. In: Proceedings of Anatidae 2000. M. Birkan, J. van Vessem, P. Havet, J. Madsen, B. Trolliet, and M. Moser (eds.
Often placed in genus Ardea.
Estimate in broad range based on size of large colonies in far N Australia and status as common and locally breeding in coastal E Australia.
Ongoing, long-term trend of increase throughout population's range.
seychellarum
2001 - 2001
1 - 10,000
1991 - 2001
Stable
WPE5
References
Dodman, T. 2002. Waterbird Population Estimates in Africa. Unpublished report to Wetlands International.
Notes
Often placed in genus Ardea.
Ardea cinerea
Grey Heron
cinerea
Sub-Saharan Africa
1995 - 2014
100,000 - 300,000
2006 - 2015
Increasing?
CSR7
References
Dodman, T. 2014. Status, Estimates and Trends of Waterbird Populations in Africa: AEWA-listed African populations. Wetlands International. (CSR6 African populations) URL: https://www.wetlands.org/publications/1304/
Wetlands International (2017) Flyway trend analyses based on data from the African-Eurasian Waterbird Census from the period of 1967-2015. Ede, The Netherlands: Wetlands International. URL: http://iwc.wetlands.org/index.php/aewatrends
Notes
approx 50,000 in Southern Africa, up to 100,000 in Eastern Africa, up to 100,000 in Western Africa, and up to 50,000 in Central Africa
Northern & Western Europe
2002 - 2013
347,000 - 712,000
2006 - 2015
Declining
CSR7
References
BirdLife International 2015. European Red List of Birds. Luxembourg: Office for Official Publications of the European Communities. http://datazone.birdlife.org/info/euroredlist
Dodman, T. 2014. Status, Estimates and Trends of Waterbird Populations in Africa: AEWA-listed African populations. Wetlands International. (CSR6 African populations) URL: https://www.wetlands.org/publications/1304/
Wetlands International (2017) Flyway trend analyses based on data from the African-Eurasian Waterbird Census from the period of 1967-2015. Ede, The Netherlands: Wetlands International. URL: http://iwc.wetlands.org/index.php/aewatrends
Notes
Total number of breeding pairs is 115,754-237,071 pairs, i.e. 347,000-711,000 individuals BirdLife International 2015). Less than 300 birds breed in North Africa (Dodman, 2014).
Moderate decline in the short-term based on breeding numbers (BirdLife International 2015) and uncertain with a negative tendency based on mid-winter counts (Wetlands International 2017). Increased in the long-term according to both sources.
Central & Eastern Europe
2000 - 2014
322,000 - 459,000
2000 - 2012
Declining
CSR7
References
BirdLife International 2015. European Red List of Birds. Luxembourg: Office for Official Publications of the European Communities. http://datazone.birdlife.org/info/euroredlist
Dodman, T. 2014. Status, Estimates and Trends of Waterbird Populations in Africa: AEWA-listed African populations. Wetlands International. (CSR6 African populations) URL: https://www.wetlands.org/publications/1304/
Wetlands International (2017) Flyway trend analyses based on data from the African-Eurasian Waterbird Census from the period of 1967-2015. Ede, The Netherlands: Wetlands International. URL: http://iwc.wetlands.org/index.php/aewatrends
Notes
In WPE2 this population belonged to one single population (E B Sea & W/SW Asia (bre)).
107,317-153,025 pairs in Europe (BirdLife International 2015). Casual breeder in Egypt (Dodman, 2014).
BirdLife International (2015) estimates the trend to be negative in the short-term and stable in the long-term. Wetlands International (2017) has also found negative short-term trend and stable long-term one.
West & South-west Asia (bre)
2000 - 2017
25,000 - 100,000
2006 - 2015
Declining?
CSR7
References
Sheldon, R. 2017. Estimates of breeding waterbird populations in Central/SW Asia, The Caucasus and the Arabian Peninsula.
Wetlands International (2017) Flyway trend analyses based on data from the African-Eurasian Waterbird Census from the period of 1967-2015. Ede, The Netherlands: Wetlands International. URL: http://iwc.wetlands.org/index.php/aewatrends
Notes
In WPE2 this population belonged to one single population (E B Sea & W/SW Asia (bre)).
See CSR6 and Sheldon (2017).
Statistically uncertain short-term trend with a negative tendency. The long-term trend is stable.
South Asia
—
100,000 - 100,000
—
Unknown
WPE5
References
Balachandran, S., in litt. 2005.
Notes
Sometimes assigned to rectirostris.
monicae
—
7,500 - 12,500
1984 - 1997
Stable
WPE5
References
Campredon, P. 1987. La reproduction des oiseaux d'eau sur le Parc National du Banc d'Arguin (Mauritanie) en 1984-1985. Alauda 55: 187-210.
Hafner, H. 2000. Herons in The Mediterranean. Pp 32-54 in: Heron Conservation (James.A. Kushlan and Heinz Hafner, Eds.). Academic Press, London.
Notes
Sometimes considered a separate species.
firasa
2001 - 2001
5,000 - 5,000
1991 - 2001
Declining
WPE5
References
Dodman, T. 2002. Waterbird Population Estimates in Africa. Unpublished report to Wetlands International.
Hawkins, F. In litt. 2002. Unpublished notes on waterbird population estimates of Madagascar.
jouyi
E, SE Asia
1987 - 1991
100,000 - 1,000,000
—
Unknown
WPE5
References
Perennou, C.P., Mundkur, T. and Scott, D.A. 1994. The Asian Waterfowl Census 1987-1991: distribution and status of Asian waterfowl. IWRB Spec. Publ. No. 24; AWB Spec. Publ. No. 86. Slimbridge, UK and Kuala Lumpur, Malaysia.
Sumatra
—
1,000 - 2,000
—
Unknown
WPE5
References
Lansdown, R.V. (pers. comm.) 1996. Data prepared for the Draft Ardeidae Action Plan.
Notes
Sometimes assigned to altirostris.
Ardea herodias
Great Blue Heron
herodias
C & NE North America
—
124,000 - 125,000
1966 - 2000
Increasing
WPE5
References
Kushlan, J. A. 2007. Conserving Herons, A Conservation Action Plan for the Herons of the World. Heron Specialist Group and Station Biologique de la Tour du Valat, Arles, France.
Sauer, J. R., J. E. Hines, and J. Fallon. 2001. The North American Breeding Bird Survey, Results and Analysis 1966 - 2000. Version 2001.2, USGS Patuxent Wildlife Research Center, Laurel, MD.
Notes
This estimate includes two other populations (W N Americas and W N Americas, N Mexico).
W North America
—
—
1966 - 2000
Stable
WPE5
References
Sauer, J. R., J. E. Hines, and J. Fallon. 2001. The North American Breeding Bird Survey, Results and Analysis 1966 - 2000. Version 2001.2, USGS Patuxent Wildlife Research Center, Laurel, MD.
Notes
Sometimes ascribed to hyperonca.
The population estimate for C & NE N America includes the estimate for this population and also the W N Americas, N Mexico population.
W North America, N Mexico
—
—
—
Unknown
WPE5
Notes
Sometimes ascribed to treganzai. (WPE4)
The population estimate for C & NE N America includes the estimate for this population and also the W N Americas population.
Baja California, Mexico
—
—
—
Unknown
WPE5
Notes
Sometimes ascribed to sanctilucae.
cognata
2004 - 2004
1,000 - 1,000
—
Unknown
WPE5
References
Boyla, K. and Estrada, A. (Eds.) 2005. ?reas Importantes para la Conservaci?n de las Aves en los Andes Tropicales: Sitios prioritarios para la conservaci?n de la biodiversidad. Quito, Ecuador: BirdLife International and Conservation International (BirdLife Conservation Series No. 14).
fannini
1997 - 2007
9,500 - 11,000
1997 - 2007
Declining
WPE5
References
COSEWIC. 2008. COSEWIC assessment and update status report on the Great Blue Heron fannini subspecies Ardea herodias fannini in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. vii + 39 pp.
Notes
Estimate refers to Nesting adults.
wardi
—
—
1966 - 2000
Increasing
WPE5
References
Sauer, J. R., J. E. Hines, and J. Fallon. 2001. The North American Breeding Bird Survey, Results and Analysis 1966 - 2000. Version 2001.2, USGS Patuxent Wildlife Research Center, Laurel, MD.
occidentalis
Florida
2006 - 2006
1,500 - 1,500
1980 - 2007
Declining
WPE5
References
Kushlan, J. A. 2007. Conserving Herons, A Conservation Action Plan for the Herons of the World. Heron Specialist Group and Station Biologique de la Tour du Valat, Arles, France.
Vennesland, R.G. and Butler, R.W. 2011. Great Blue Heron (Ardea herodias), The Birds of North America Online (A. Poole, Ed.). Ithaca: Cornell Lab of Ornithology.
Caribbean
—
—
—
Unknown
WPE5
Notes
According to Birdlife International 2008a., occurs mainly as non-breeding migrant. Resident population presumably <100 individuals.
Central America
2005 - 2005
29,800 - 122,000
—
Unknown
WPE5
References
Komar, O., Angehr, G., Eisermann, K., Herrera, N. and Zolotoff, J.M.. 2006. Waterbird Population Estimates for Central America. IV North American Ornithological Conference. unpublished report.
Ardea cocoi
Cocoi Heron
South America, Panama
2005 - 2011
100,000 - 1,000,000
2001 - 2010
Stable
WPE5
References
Huanca Llanos, N.E. 2006. Reporte final, Aves Acu?ticas en Bolivia. Asociaci?n Armonia, Santa Cruz de la Sierra (unpublished report to BirdLife International).
Ottema, O. 2006. Waterbirds in Suriname: Final report. STINASU Foundation for Nature Conservation Suriname, Paramaribo (unpublished report).
Santander, T., J.R. Hidálgo and B. Haase. 2006. Reporte final Aves Acuáticas en Ecuador. Aves & Conservación, Quito (unpublished report).
Unterkofler, D. and Blanco, D.E. 2012. Unpublished information on Argentine waterbirds.
Wetlands International Argentina (compiler). 2012. Unpublished information from the Neotropical Waterbird Census, 1990-2011, including contributions from Colombia (Asociaci?n Calidris in litt.), Paraguay (Guyra Paraguay in litt.), Uruguay (Averaves in litt.) and Venezuela (Uni?n Venezolana de Ornit?logos in litt.).
Ardea pacifica
White-necked Heron
Australia
2004 - 2011
10,000 - 25,000
1982 - 2011
Fluctuating
WPE5
References
Jaensch, Roger, August 2005. Detailed data and notes on numbers and distribution of waterbirds in Australia, updated from Jaensch, 2003, and derived from more than 90 Australian sources including Wetlands International, Birds Australia and Jaensch, 2003,
Jaensch, R. April 2012. Detailed data and notes on numbers and trends of waterbirds in Australia and other parts of the Oceania region, in a spreadsheet prepared for Wetlands International - Oceania for the 5th edition of WPE.
Notes
Estimate based on reassessment of previous information and recent information that suggests population level may reach or exceed 25,000 only briefly.
Ardea melanocephala
Black-headed Heron
Sub-Saharan Africa
1991 - 2001
100,000 - 500,000
2006 - 2015
Unknown
CSR7
References
Dodman, T. 2014. Status, Estimates and Trends of Waterbird Populations in Africa: AEWA-listed African populations. Wetlands International. (CSR6 African populations) URL: https://www.wetlands.org/publications/1304/
Wetlands International (2017) Flyway trend analyses based on data from the African-Eurasian Waterbird Census from the period of 1967-2015. Ede, The Netherlands: Wetlands International. URL: http://iwc.wetlands.org/index.php/aewatrends
Ardea humbloti
Madagascar Heron
Madagascar
—
1,000 - 3,000
1990 - 2005
Declining
WPE5
References
BirdLife International. 2005. Threatened birds of the world 2005. Species factsheets available at?www.birdlife.org
Hawkins, F. In litt. 2002. Unpublished notes on waterbird population estimates of Madagascar.
Ardea insignis
White-bellied Heron
S & SE Asia
2005 - 2005
250 - 1,000
1985 - 2005
Declining
WPE5
References
BirdLife International. 2005. Threatened birds of the world 2005. Species factsheets available at?www.birdlife.org
Unpublished information supplied by Wetlands International Specialist Groups, 2006.
Ardea sumatrana
Great-billed Heron
SE Asia
—
10,000 - 100,000
1975 - 1985
Declining
WPE5
References
Lansdown, R.V. 1986. The status and ecology of the Sumatran Heron (Ardea sumatrana). Interwader publ. No. 21. Kuala Lumpur.
Australia
1999 - 1999
7,500 - 7,500
1990 - 1999
Stable
WPE5
References
Green, A.J. 1992a. The status and conservation of the White-winged Wood Duck (Cairina scutulata). IWRB Special Publication No. 17. Slimbridge, UK. 115 pp.
Notes
Sometimes ascribed to mathewsae.
Occurs at low density often in habitat remote from observation; thus, data deficient. Habitat is largely unchanged and not at high risk.
Ardea goliath
Goliath Heron
Sub-Saharan Africa
—
10,000 - 100,000
1991 - 2001
Stable
WPE5
References
del Hoyo, J., Elliott A. and Sargatal, J. (eds). 1992. Handbook of the Birds of the World. Volume 1: Ostrich to Ducks. Lynx Editions, Barcelona.
Dodman, T. 2002. Waterbird Population Estimates in Africa. Unpublished report to Wetlands International.
SW Asia
1987 - 1991
50 - 50
—
Unknown
WPE5
References
Perennou, C.P., Mundkur, T. and Scott, D.A. 1994. The Asian Waterfowl Census 1987-1991: distribution and status of Asian waterfowl. IWRB Spec. Publ. No. 24; AWB Spec. Publ. No. 86. Slimbridge, UK and Kuala Lumpur, Malaysia.
S Asia
1987 - 1991
20 - 20
—
Unknown
WPE5
References
Perennou, C.P., Mundkur, T. and Scott, D.A. 1994. The Asian Waterfowl Census 1987-1991: distribution and status of Asian waterfowl. IWRB Spec. Publ. No. 24; AWB Spec. Publ. No. 86. Slimbridge, UK and Kuala Lumpur, Malaysia.
Ardea purpurea
Purple Heron
purpurea
Tropical Africa
2001 - 2001
75,000 - 100,000
2005 - 2015
Declining?
CSR7
References
Dodman, T. 2014. Status, Estimates and Trends of Waterbird Populations in Africa: AEWA-listed African populations. Wetlands International. (CSR6 African populations) URL: https://www.wetlands.org/publications/1304/
Wetlands International (2017) Flyway trend analyses based on data from the African-Eurasian Waterbird Census from the period of 1967-2015. Ede, The Netherlands: Wetlands International. URL: http://iwc.wetlands.org/index.php/aewatrends
Notes
According to the IWC trend analysis data, the population is possibly in significant long-term decline although only partial information is available (Wetlands International 2017).
West Europe & West Mediterranean/West Africa
2000 - 2012
32,000 - 38,000
2000 - 2012
Declining
CSR7
References
BirdLife International 2015. European Red List of Birds. Luxembourg: Office for Official Publications of the European Communities. http://datazone.birdlife.org/info/euroredlist
Dodman, T. 2014. Status, Estimates and Trends of Waterbird Populations in Africa: AEWA-listed African populations. Wetlands International. (CSR6 African populations) URL: https://www.wetlands.org/publications/1304/
Notes
10,802-12.400 pairs in CH, NL, DE, IT, FR, ES and PT (BirdLife International 2015). Less than 300 in North Africa (Dodman, 2014).
Declining in the short-term. Long-term trend appears to be stable (BirdLife International 2015).
East Europe, Black Sea & Mediterranean/Sub-Saharan Africa
2000 - 2013
61,000 - 99,000
2006 - 2015
Declining?
CSR7
References
BirdLife International 2015. European Red List of Birds. Luxembourg: Office for Official Publications of the European Communities. http://datazone.birdlife.org/info/euroredlist
Dodman, T. 2014. Status, Estimates and Trends of Waterbird Populations in Africa: AEWA-listed African populations. Wetlands International. (CSR6 African populations) URL: https://www.wetlands.org/publications/1304/
Notes
20,411-32,945 pairs (BirdLife International 2015). This estimate is without the estimate for SW Asian part of the population, which was split from.
Declining both in the short- and the long-term. Unknown breeding trends are reported from 7 of the 20 breeding range states (BirdLife International 2015).
SW Asia (bre)
2006 - 2006
10,000 - 25,000
2006 - 2015
Unknown
CSR7
References
Wetlands International (2017) Flyway trend analyses based on data from the African-Eurasian Waterbird Census from the period of 1967-2015. Ede, The Netherlands: Wetlands International. URL: http://iwc.wetlands.org/index.php/aewatrends
Notes
In WPE2 this population belonged to one single population (E Europe/SW Asia (breeding)).
WI/IUCN Heron SG (2005).
madagascariensis
—
5,000 - 10,000
1991 - 2001
Stable
WPE5
References
Dodman, T. 2002. Waterbird Population Estimates in Africa. Unpublished report to Wetlands International.
Hawkins, F. In litt. 2002. Unpublished notes on waterbird population estimates of Madagascar.
bournei
—
1 - 50
1982 - 1992
Declining
WPE5
References
Hazevoet, C.J. 1992. A review of the Santiago Purple Heron (Ardea purpurea bournei) with a report of a new colony. Bird Conservation International (1992) 2: 15-23.
Notes
May merit full specific status (del Hoyo et al. (1992)).
manilensis
South Asia
1987 - 1991
25,000 - 25,000
1977 - 1991
Stable
WPE5
References
Perennou, C.P., Mundkur, T. and Scott, D.A. 1994. The Asian Waterfowl Census 1987-1991: distribution and status of Asian waterfowl. IWRB Spec. Publ. No. 24; AWB Spec. Publ. No. 86. Slimbridge, UK and Kuala Lumpur, Malaysia.
E & SE Asia
1987 - 1991
10,000 - 100,000
—
Unknown
WPE5
References
Perennou, C.P., Mundkur, T. and Scott, D.A. 1994. The Asian Waterfowl Census 1987-1991: distribution and status of Asian waterfowl. IWRB Spec. Publ. No. 24; AWB Spec. Publ. No. 86. Slimbridge, UK and Kuala Lumpur, Malaysia.
Ardea alba
Great White Egret
alba
W, C & SE Europe/Black Sea & Mediterranean
2000 - 2014
61,000 - 99,000
2006 - 2015
Stable/ Increasing?
CSR7
References
BirdLife International 2015. European Red List of Birds. Luxembourg: Office for Official Publications of the European Communities. http://datazone.birdlife.org/info/euroredlist
Wetlands International (2017) Flyway trend analyses based on data from the African-Eurasian Waterbird Census from the period of 1967-2015. Ede, The Netherlands: Wetlands International. URL: http://iwc.wetlands.org/index.php/aewatrends
Notes
Often assigned to genus Casmerodius, occasionally Egretta.
20,248-32,928 pairs in Europe (BirdLife International 2015). Possibly, some birds in the Volga delta belong to the Western Asia/South-west Asia population.
In the short-term, increased based on breeding numbers (BirdLife International 2015), but stabilized based on wintering numbers (Wetlands International 2017). In the long-term, increase based on both source.
Western Asia/South-west Asia
1990 - 2017
25,000 - 100,000
2006 - 2015
Stable/ Fluctuating
CSR7
References
Wetlands International (2017) Flyway trend analyses based on data from the African-Eurasian Waterbird Census from the period of 1967-2015. Ede, The Netherlands: Wetlands International. URL: http://iwc.wetlands.org/index.php/aewatrends
Notes
See CSR7 and Sheldon (2017).
Stable both in the short- and the long-term (Wetlands International 2017). The trend graph shows increase up to the late 1990s, followed by a rapid decline in the early 2000s and stabilisation in the last decade.
melanorhynchos
Sub-Saharan Africa & Madagascar
2001 - 2001
100,000 - 500,000
2006 - 2015
Stable
CSR7
References
Dodman, T. 2002. Waterbird Population Estimates in Africa. Unpublished report to Wetlands International.
Wetlands International (2017) Flyway trend analyses based on data from the African-Eurasian Waterbird Census from the period of 1967-2015. Ede, The Netherlands: Wetlands International. URL: http://iwc.wetlands.org/index.php/aewatrends
Notes
Notes
Stable both in the short- and the long-term.
egretta
North America
—
270,000 - 270,000
1966 - 2000
Increasing
WPE5
References
Kushlan, James, A., Melanie J. Steinkamp, Katharine Parsons, Jack Capp, Martin Acosta Cruz, Malcolm Coulter, Ian Davidson, Loney Dickson, Naomi Edelson, Richard Elliot, R. Michael Erwin, Scott Hatch, Stephen Kress, Robert Milko, Steve Miller, Kyra Mills, Richard Paul, Roberto Phillips, Jorge E. Saliva, Bill Sydeman, John Trapp, Jennifer Wheeler, and Kent Wohl. 2002. North American Waterbird Conservation Plan, Version 1. Waterbird Conservation for the Americas, Washington, DC, USA.
Sauer, J. R., J. E. Hines, and J. Fallon. 2001. The North American Breeding Bird Survey, Results and Analysis 1966 - 2000. Version 2001.2, USGS Patuxent Wildlife Research Center, Laurel, MD.
Mexico
—
—
—
Unknown
WPE5
Notes
Population added in WPE3.
Central America
2005 - 2005
52,500 - 180,000
2005 - 2005
Stable?
WPE5
References
Komar, O., Angehr, G., Eisermann, K., Herrera, N. and Zolotoff, J.M.. 2006. Waterbird Population Estimates for Central America. IV North American Ornithological Conference. unpublished report.
Total of data presented in BirdLife International 2008a.
South America
2005 - 2011
1,000,000 - 1,000,000
2001 - 2010
Stable
WPE5
References
Huanca Llanos, N.E. 2006. Reporte final, Aves Acu?ticas en Bolivia. Asociaci?n Armonia, Santa Cruz de la Sierra (unpublished report to BirdLife International).
Ottema, O. 2006. Waterbirds in Suriname: Final report. STINASU Foundation for Nature Conservation Suriname, Paramaribo (unpublished report).
Santander, T., J.R. Hidálgo and B. Haase. 2006. Reporte final Aves Acuáticas en Ecuador. Aves & Conservación, Quito (unpublished report).
Unterkofler, D. and Blanco, D.E. 2012. Unpublished information on Argentine waterbirds.
Wetlands International Argentina (compiler). 2012. Unpublished information from the Neotropical Waterbird Census, 1990-2011, including contributions from Colombia (Asociaci?n Calidris in litt.), Paraguay (Guyra Paraguay in litt.), Uruguay (Averaves in litt.) and Venezuela (Uni?n Venezolana de Ornit?logos in litt.).
Notes
Population added in WPE3.
modestus
South Asia (non-bre)
2001 - 2005
25,000 - 100,000
1977 - 1991
Stable
WPE5
References
Asian Waterbird Census unpublished data. 2002.
Perennou, C.P., Mundkur, T. and Scott, D.A. 1994. The Asian Waterfowl Census 1987-1991: distribution and status of Asian waterfowl. IWRB Spec. Publ. No. 24; AWB Spec. Publ. No. 86. Slimbridge, UK and Kuala Lumpur, Malaysia.
ul Islam, Zafar, in litt. 2005.
Notes
Sometimes considered as Ardea (Casmerodius, Egretta) modestus.
20: 11,000 counted in South Asia in 2003.
E Asia (non-bre)
2001 - 2001
10,000 - 100,000
1987 - 1991
Declining
WPE5
References
Asian Waterbird Census unpublished data. 2002.
Perennou, C.P., Mundkur, T. and Scott, D.A. 1994. The Asian Waterfowl Census 1987-1991: distribution and status of Asian waterfowl. IWRB Spec. Publ. No. 24; AWB Spec. Publ. No. 86. Slimbridge, UK and Kuala Lumpur, Malaysia.
Notes
Sometimes considered as Ardea (Casmerodius, Egretta) modestus. Two forms are recognised by ornithologists in the region (one closely resembling alba) and research is needed on their taxonomy and distribution (Butler et al. (1992), Moores, N. in litt. (2005)).
20: 16,800 counted in 2004.
Indonesia
—
—
—
Unknown
WPE5
Notes
Sometimes considered as Ardea (Casmerodius, Egretta) modestus.
Australia
1995 - 2011
25,000 - 100,000
1982 - 2011
Fluctuating
WPE5
References
Jaensch, Roger, August 2005. Detailed data and notes on numbers and distribution of waterbirds in Australia, updated from Jaensch, 2003, and derived from more than 90 Australian sources including Wetlands International, Birds Australia and Jaensch, 2003,
Jaensch, R. April 2012. Detailed data and notes on numbers and trends of waterbirds in Australia and other parts of the Oceania region, in a spreadsheet prepared for Wetlands International - Oceania for the 5th edition of WPE.
Halse, S.A., Pearson, G.B., Jaensch, R.P., Kulmoi, P., Gregory, P., Kay, W.R. and Storey, A.W. 1996. Waterbird surveys of the Middle Fly River floodplain, Papua New Guinea. Wildlife Research 23 (5): 557-569. (CSIRO Publishing, Australia).
Notes
Sometimes considered as Ardea (Casmerodius, Egretta) modestus.
Estimate based on review of previous information and recent information that suggests total numbers remain between 25,000 and 100,000. Numbers in New Guinea can reach the same level but likely to be mostly migrants from Australia.
New Zealand
—
100 - 100
—
Unknown
WPE5
References
Powlesland, R. (pers. comm.). 1993. New Zealand Department of Conservation unpublished data.
Notes
Sometimes considered as Ardea (Casmerodius, Egretta) modestus. Sometimes ascribed to maorianus.
Ardea brachyrhyncha
Yellow-billed Egret
Sub-Saharan Africa
2001 - 2001
25,000 - 100,000
2006 - 2015
Increasing?
CSR7
References
Dodman, T. 2002. Waterbird Population Estimates in Africa. Unpublished report to Wetlands International.
Wetlands International (2017) Flyway trend analyses based on data from the African-Eurasian Waterbird Census from the period of 1967-2015. Ede, The Netherlands: Wetlands International. URL: http://iwc.wetlands.org/index.php/aewatrends
Notes
Ardea intermedia, A. brachyrhyncha and A. plumifera (del Hoyo and Collar 2014) were previously placed in the genus Mesophoyx and lumped as M. intermedia (see BirdLife International (2016) Species factsheet: Ardea brachyrhyncha.)
Ardea intermedia
Intermediate Egret
intermedia
South Asia
—
25,000 - 100,000
—
Unknown
WPE5
References
Asian Waterbird Census unpublished data. 2002.
Notes
Often included in the genus Mesophoyx or Egretta.
20: 22,000 counted in South Asia in 2004.
E, SE Asia
—
25,000 - 100,000
—
Unknown
WPE5
References
Asian Waterbird Census unpublished data. 2002.
Notes
20: >10,000 counted in 2002-2004.
Ardea plumifera
Plumed Egret
plumifera
1995 - 2011
100,000 - 1,000,000
1982 - 2011
Declining?
WPE5
References
BirdLife International. 2012h. Important Bird Areas factsheet: Gwydir Wetlands. Downloaded from?http://www.birdlife.org?on 30/04/2012
Halse, S.A., Pearson, G.B., Jaensch, R.P., Kulmoi, P., Gregory, P., Kay, W.R. and Storey, A.W. 1996. Waterbird surveys of the Middle Fly River floodplain, Papua New Guinea. Wildlife Research 23 (5): 557-569. (CSIRO Publishing, Australia).
Jaensch, Roger, August 2005. Detailed data and notes on numbers and distribution of waterbirds in Australia, updated from Jaensch, 2003, and derived from more than 90 Australian sources including Wetlands International, Birds Australia and Jaensch, 2003,
Jaensch, R. April 2012. Detailed data and notes on numbers and trends of waterbirds in Australia and other parts of the Oceania region, in a spreadsheet prepared for Wetlands International - Oceania for the 5th edition of WPE.
Kingsford, R.T. and Johnson, W. 1998. Impact of water diversions on colonially-nesting waterbirds in the Macquarie Marshes of arid Australia. Colonial Waterbirds 21: 159-170.
Kingsford, R.T., Porter, J.L. and Halse, S.A. 2012. National waterbird assessment. Waterlines report, National Water Commission, Canberra. Available online at http://nwc.gov.au/publications/waterlines/74
Notes
Ardea intermedia, A. brachyrhyncha and A. plumifera (del Hoyo and Collar 2014) were previously placed in the genus Mesophoyx and lumped as M. intermedia (see BirdLife International (2016) Species factsheet: Ardea plumifera.)
Numbers clearly between 100,000 and 1,000,000 either in Australia or S New Guinea; probable migration precludes summation. Additional birds in Indonesia (no information) or failure to confirm migration would raise possibility of estimate in excess of 1,000,000.
In Australia, northern colonies secure but overall likely declining trend due to loss of southernmost colonies due to river regulation for irrigation.
Syrigma sibilatrix
Whistling Heron
sibilatrix
2005 - 2011
25,000 - 100,000
2001 - 2010
Stable
WPE5
References
Huanca Llanos, N.E. 2006. Reporte final, Aves Acu?ticas en Bolivia. Asociaci?n Armonia, Santa Cruz de la Sierra (unpublished report to BirdLife International).
Unterkofler, D. and Blanco, D.E. 2012. Unpublished information on Argentine waterbirds.
Wetlands International Argentina (compiler). 2012. Unpublished information from the Neotropical Waterbird Census, 1990-2011, including contributions from Colombia (Asociaci?n Calidris in litt.), Paraguay (Guyra Paraguay in litt.), Uruguay (Averaves in litt.) and Venezuela (Uni?n Venezolana de Ornit?logos in litt.).
forstersmithi
2011 - 2011
10,000 - 10,000
2006 - 2010
Fluctuating
WPE5
References
Uni?n Venezolana de Ornit?logos. 2012. Unpublished information from the Neotropical Waterbird Census in Venezuela, 2006-2011.
Pilherodius pileatus
Capped Heron
C & S America
2005 - 2011
10,000 - 25,000
2001 - 2010
Stable
WPE5
References
Uni?n Venezolana de Ornit?logos. 2012. Unpublished information from the Neotropical Waterbird Census in Venezuela, 2006-2011.
Egretta picata
Pied Heron
Australia - Sulawesi
1995 - 2002
25,000 - 100,000
2000 - 2010
Stable
WPE5
References
Bishop, K.D. 2003. A Review of the Avifauna of the TransFly Eco-region: the status, distribution, habitats and conservation of the region?s birds. Report to WWF South Pacific Program.
Chatto, R. 2000. Waterbird breeding colonies in the Top End of the Northern Territory. Technical Report 69, Parks and Wildlife Commission of the Northern Territory, Australia.
Halse, S.A., Pearson, G.B., Jaensch, R.P., Kulmoi, P., Gregory, P., Kay, W.R. and Storey, A.W. 1996. Waterbird surveys of the Middle Fly River floodplain, Papua New Guinea. Wildlife Research 23 (5): 557-569. (CSIRO Publishing, Australia).
Jaensch, Roger, August 2005. Detailed data and notes on numbers and distribution of waterbirds in Australia, updated from Jaensch, 2003, and derived from more than 90 Australian sources including Wetlands International, Birds Australia and Jaensch, 2003,
Jaensch, R. April 2012. Detailed data and notes on numbers and trends of waterbirds in Australia and other parts of the Oceania region, in a spreadsheet prepared for Wetlands International - Oceania for the 5th edition of WPE.
Notes
Sometimes placed in genus Ardea.
Estimate based on high counts in S New Guinea, and numbers breeding in far N Australia, which although not necessarily additive nevertheless suggest overall numbers in the range 25,000-100,000.
Trend assumed stable based on habitat in Australia and New Guinea being mostly near-natural.
Egretta novaehollandiae
White-faced Heron
novaehollandiae
2004 - 2008
10,000 - 100,000
—
Unknown
WPE5
References
Jaensch, Roger, August 2005. Detailed data and notes on numbers and distribution of waterbirds in Australia, updated from Jaensch, 2003, and derived from more than 90 Australian sources including Wetlands International, Birds Australia and Jaensch, 2003,
Joseph, L. In litt. 4 April 2012 (CSIRO, Canberra).
Madsen, J., Reed, A. and Andreev, A. 1996. Status and trends of geese (Anser sp., Branta sp.) in the world: a review, updating and evaluation. In: Proceedings of Anatidae 2000. M. Birkan, J. van Vessem, P. Havet, J. Madsen, B. Trolliet, and M. Moser (eds.
Reid, J.R.W., Kingsford, R.T. and Jaensch, R.P. 2009. Waterbird Surveys in the Channel Country Floodplain Wetlands, Autumn 2009. Report by Australian National University, Canberra, University of New South Wales, Sydney, and Wetlands International, Oceania, Brisbane, for the Australian Government Department of Environment, Water, Heritage and the Arts. 81 pp. Also available online at http://www.lebmf.gov.au/publications/pubs/waterbirds-report.pdf
Notes
Considered to be monotypic by Marchant & Higgins (1990) & Boles & Walter, in litt. (2005).
Estimate based on summed maximum counts indicating at least 10,000 birds and common status (at low density) across dryland habitats; much habitat is not surveyed.
parryi
—
—
—
Unknown
WPE5
Notes
Population added in WPE3.
Egretta rufescens
Reddish Egret
rufescens
S USA
1991 - 2006
5,000 - 7,000
1991 - 2006
Declining
WPE5
References
Green, M.C. 2006. Status report and survey recommendations on the Reddish Egret (Egretta rufescens). U.S. Fish and Wildlife Service, Atlanta, GA. 37 pp.
Notes
In WPE2 this population belonged to one single population (S.h. rufescens, North America/Caribbean).
Green 2006. 1,215-1,330 breeding pairs in southern USA; 800-1,000 breeding pairs in Mexico (inc. dickeyi); 200 breeding pairs in Bahamas.
Mexico
—
1 - 10,000
—
Unknown
WPE5
References
Scott, D.A. and Carbonnell, M. (compilers). 1986. A Directory of Neotropical Wetlands. IUCN, Cambridge, UK and IWRB, Slimbridge, UK.
Notes
Sometimes ascribed to colorata.
Central America
2006 - 2006
1 - 10,000
—
Unknown
WPE5
References
Unpublished information supplied by Wetlands International Specialist Groups, 2006.
Notes
In WPE2 this population belonged to one single population (S.h. rufescens, North America/Caribbean).
Green, C. In litt. 2012. 282 breeding pairs in southern Mexico (Chiapas, Oaxaca in 2007-2008) and 95 breeding pairs in Tamaulipas (in 2008).
Caribbean
2006 - 2010
213 - 213
—
Unknown
WPE5
References
Kushlan, J. A. 2007. Conserving Herons, A Conservation Action Plan for the Herons of the World. Heron Specialist Group and Station Biologique de la Tour du Valat, Arles, France.
Notes
In WPE2 this population belonged to one single population (S.h. rufescens, North America/Caribbean).
Data from Bahamas: 50 bp in Great Inagua, 19 bp in Grand Bahama, 2 bp in Bimini Island.
N South America
—
1 - 10,000
—
Unknown
WPE5
References
Morales, G. 2000. Herons in South America. Pp 177-199 in: Heron Conservation (James.A. Kushlan and Heinz Hafner, Eds.). Academic Press, London.
Notes
In WPE2 this population belonged to one single population (S.h. rufescens, North America/Caribbean).
dickeyi
2007 - 2008
2,250 - 2,260
—
Unknown
WPE5
References
Green, C. In litt. 2012.
Notes
753 Breeding Pairs.
Egretta ardesiaca
Black Heron
Sub-Saharan Africa
1999 - 1999
25,000 - 100,000
2006 - 2015
Unknown
CSR7
References
del Hoyo, J., Elliott A. and Sargatal, J. (eds). 1992. Handbook of the Birds of the World. Volume 1: Ostrich to Ducks. Lynx Editions, Barcelona.
Wetlands International (2017) Flyway trend analyses based on data from the African-Eurasian Waterbird Census from the period of 1967-2015. Ede, The Netherlands: Wetlands International. URL: http://iwc.wetlands.org/index.php/aewatrends
Egretta vinaceigula
Slaty Egret
Central Southern Africa
2005 - 2005
3,000 - 5,000
1993 - 2013
Declining?
CSR7
References
BirdLife International (2014) Species factsheet: Egretta vinaceigula. Downloaded from http://www.birdlife.org on 07/10/2014. Recommended citation for factsheets for more than one species: BirdLife International (2014) IUCN Red List for birds. Downloaded from http://www.birdlife.org on 07/10/2014.
BirdLife International (2017) IUCN Red List for birds. Downloaded from http://www.birdlife.org on 30/09/2017.
Wetlands International (2017) Flyway trend analyses based on data from the African-Eurasian Waterbird Census from the period of 1967-2015. Ede, The Netherlands: Wetlands International. URL: http://iwc.wetlands.org/index.php/aewatrends
Notes
No update to estimate in AEWA SSAP (Tyler 2013)
The population is suspected to be in decline owing to the effects of habitat conversion and degradation, and human disturbance. The likely rate of decline, however, has not been estimated (BirdLife International, 2017). Recent IWC trend analysis provides some week support to this assumption (Wetlands International 2017). Significant long-term decline maintained.
Egretta tricolor
Tricolored Heron
tricolor
—
—
—
Unknown
WPE5
ruficollis
USA (bre)
—
293,000 - 293,000
1966 - 2000
Stable
WPE5
References
Kushlan, James, A., Melanie J. Steinkamp, Katharine Parsons, Jack Capp, Martin Acosta Cruz, Malcolm Coulter, Ian Davidson, Loney Dickson, Naomi Edelson, Richard Elliot, R. Michael Erwin, Scott Hatch, Stephen Kress, Robert Milko, Steve Miller, Kyra Mills, Richard Paul, Roberto Phillips, Jorge E. Saliva, Bill Sydeman, John Trapp, Jennifer Wheeler, and Kent Wohl. 2002. North American Waterbird Conservation Plan, Version 1. Waterbird Conservation for the Americas, Washington, DC, USA.
Sauer, J. R., J. E. Hines, and J. Fallon. 2001. The North American Breeding Bird Survey, Results and Analysis 1966 - 2000. Version 2001.2, USGS Patuxent Wildlife Research Center, Laurel, MD.
Notes
In WPE2 this population belonged to one single population (E.t. ruficollis, North Am/NW Neotropics).
Mexico
2006 - 2006
1 - 1,500
—
Unknown
WPE5
References
Kushlan, J. A. 2007. Conserving Herons, A Conservation Action Plan for the Herons of the World. Heron Specialist Group and Station Biologique de la Tour du Valat, Arles, France.
Notes
Sometimes ascribed to rufimentum.
No new data
Central America
2005 - 2005
3,350 - 6,800
—
Unknown
WPE5
References
Komar, O., Angehr, G., Eisermann, K., Herrera, N. and Zolotoff, J.M.. 2006. Waterbird Population Estimates for Central America. IV North American Ornithological Conference. unpublished report.
Notes
In WPE2 this population belonged to one single population (E.t. ruficollis, North Am/NW Neotropics).
In WPE2 this population belonged to one single population (E.t. ruficollis, North Am/NW Neotropics).
Total of data presented in BirdLife International 2008a and of which 10,000-25,000 estimated for Cuba (Acosta-Cruz, and Mugica-Vald?s 2006).
Egretta caerulea
Little Blue Heron
North America
—
225,000 - 300,000
1966 - 2000
Declining
WPE5
References
Butler, R.W., Kushlan, J.A. and Davidson, I.J. 2000. Herons in North America, Central America and the West Indies. Pp 151-175 in: Heron Conservation (James.A. Kushlan and Heinz Hafner, Eds.). Academic Press, London.
Sauer, J. R., J. E. Hines, and J. Fallon. 2001. The North American Breeding Bird Survey, Results and Analysis 1966 - 2000. Version 2001.2, USGS Patuxent Wildlife Research Center, Laurel, MD.
Notes
In WPE2 this population belonged to one single population (North America/Neotropics).
Mexico
—
75,000 - 150,000
—
Unknown
WPE5
References
Butler, R.W., Kushlan, J.A. and Davidson, I.J. 2000. Herons in North America, Central America and the West Indies. Pp 151-175 in: Heron Conservation (James.A. Kushlan and Heinz Hafner, Eds.). Academic Press, London.
Notes
In WPE2 this population belonged to one single population (North America/Neotropics).
Central America
2005 - 2005
701 - 1,500
—
Unknown
WPE5
References
Komar, O., Angehr, G., Eisermann, K., Herrera, N. and Zolotoff, J.M.. 2006. Waterbird Population Estimates for Central America. IV North American Ornithological Conference. unpublished report.
Notes
In WPE2 this population belonged to one single population (North America/Neotropics).
In WPE2 this population belonged to one single population (North America/Neotropics).
Total of data presented in BirdLife International 2008a.
South America
2005 - 2011
100,000 - 100,000
2001 - 2010
Stable?
WPE5
References
Ottema, O. 2006. Waterbirds in Suriname: Final report. STINASU Foundation for Nature Conservation Suriname, Paramaribo (unpublished report).
Santander, T., J.R. Hidálgo and B. Haase. 2006. Reporte final Aves Acuáticas en Ecuador. Aves & Conservación, Quito (unpublished report).
Unterkofler, D. and Blanco, D.E. 2012. Unpublished information on Argentine waterbirds.
Wetlands International Argentina (compiler). 2012. Unpublished information from the Neotropical Waterbird Census, 1990-2011, including contributions from Colombia (Asociaci?n Calidris in litt.), Paraguay (Guyra Paraguay in litt.), Uruguay (Averaves in litt.) and Venezuela (Uni?n Venezolana de Ornit?logos in litt.).
Notes
In WPE2 this population belonged to one single population (North America/Neotropics).
Egretta thula
Snowy Egret
thula
—
215,000 - 215,000
1982 - 1992
Stable
WPE5
References
Canevari, P. (pers. comm.). 1993. Unpublished information from Neotropical Wetlands Program.
Kushlan, James, A., Melanie J. Steinkamp, Katharine Parsons, Jack Capp, Martin Acosta Cruz, Malcolm Coulter, Ian Davidson, Loney Dickson, Naomi Edelson, Richard Elliot, R. Michael Erwin, Scott Hatch, Stephen Kress, Robert Milko, Steve Miller, Kyra Mills, Richard Paul, Roberto Phillips, Jorge E. Saliva, Bill Sydeman, John Trapp, Jennifer Wheeler, and Kent Wohl. 2002. North American Waterbird Conservation Plan, Version 1. Waterbird Conservation for the Americas, Washington, DC, USA.
Notes
In WPE2 this population belonged to one single population (E.t. thula, Neotropics).
thula
Mexico
—
75,000 - 150,000
—
Unknown
WPE5
References
Butler, R.W., Kushlan, J.A. and Davidson, I.J. 2000. Herons in North America, Central America and the West Indies. Pp 151-175 in: Heron Conservation (James.A. Kushlan and Heinz Hafner, Eds.). Academic Press, London.
Notes
In WPE2 this population belonged to one single population (E.t. thula, Neotropics).
Central America
2005 - 2005
11,300 - 26,900
—
Unknown
WPE5
References
Komar, O., Angehr, G., Eisermann, K., Herrera, N. and Zolotoff, J.M.. 2006. Waterbird Population Estimates for Central America. IV North American Ornithological Conference. unpublished report.
Notes
In WPE2 this population belonged to one single population (E.t. thula, Neotropics).
In WPE2 this population belonged to one single population (E.t. thula, Neotropics).
Total of data presented in BirdLife International 2008a.
South America
2005 - 2011
300,000 - 1,000,000
2001 - 2010
Stable
WPE5
References
Huanca Llanos, N.E. 2006. Reporte final, Aves Acu?ticas en Bolivia. Asociaci?n Armonia, Santa Cruz de la Sierra (unpublished report to BirdLife International).
Ottema, O. 2006. Waterbirds in Suriname: Final report. STINASU Foundation for Nature Conservation Suriname, Paramaribo (unpublished report).
Wetlands International Argentina (compiler). 2012. Unpublished information from the Neotropical Waterbird Census, 1990-2011, including contributions from Colombia (Asociaci?n Calidris in litt.), Paraguay (Guyra Paraguay in litt.), Uruguay (Averaves in litt.) and Venezuela (Uni?n Venezolana de Ornit?logos in litt.).
Notes
In WPE2 this population belonged to one single population (E.t. thula, Neotropics).
brewsteri
2006 - 2006
100,000 - 1,000,000
1966 - 2000
Increasing
WPE5
References
Sauer, J. R., J. E. Hines, and J. Fallon. 2001. The North American Breeding Bird Survey, Results and Analysis 1966 - 2000. Version 2001.2, USGS Patuxent Wildlife Research Center, Laurel, MD.
Unpublished information supplied by Wetlands International Specialist Groups, 2006.
Egretta garzetta
Little Egret
garzetta
Sub-Saharan Africa
2001 - 2001
200,000 - 500,000
2006 - 2015
Increasing?
CSR7
References
Dodman, T. 2002. Waterbird Population Estimates in Africa. Unpublished report to Wetlands International.
Wetlands International (2017) Flyway trend analyses based on data from the African-Eurasian Waterbird Census from the period of 1967-2015. Ede, The Netherlands: Wetlands International. URL: http://iwc.wetlands.org/index.php/aewatrends
Notes
Trend analyses based on IWC July data suggest a significant increase, however data are rather limited to a few key countries.
Western Europe, NW Africa
2002 - 2013
106,000 - 116,000
2000 - 2012
Declining
CSR7
References
BirdLife International 2015. European Red List of Birds. Luxembourg: Office for Official Publications of the European Communities. http://datazone.birdlife.org/info/euroredlist
Dodman, T. 2014. Status, Estimates and Trends of Waterbird Populations in Africa: AEWA-listed African populations. Wetlands International. (CSR6 African populations) URL: https://www.wetlands.org/publications/1304/
Notes
Population was omitted from WPE2.
34,668-34,472 pairs in BE, ES, ESIC, FR, IE, IT, NL, PT & UK (BirdLife International 2015). 1500-3500 resident birds can be also added for NW Africa (Dodman, 2014).
Declines in the short-term but increased in the long one.
Central & E Europe, Black Sea, E Mediterranean
2000 - 2014
60,000 - 89,000
2000 - 2012
Stable
CSR7
References
BirdLife International 2015. European Red List of Birds. Luxembourg: Office for Official Publications of the European Communities. http://datazone.birdlife.org/info/euroredlist
Dodman, T. 2014. Status, Estimates and Trends of Waterbird Populations in Africa: AEWA-listed African populations. Wetlands International. (CSR6 African populations) URL: https://www.wetlands.org/publications/1304/
Notes
19,598-29,059 pairs in AL, AT, BA, BG, CY, CZ, GE, GR, HR, HU, MD, ME, MK, PL, RO, RS, RU, SK, TR, UA & XK (BirdLife International 2015) allocating 40% of the Russian population to this one. According to Dodman (2014), further 1000-2000 resident birds can be added for Egypt.
Stable in the short-term and stable/fluctuating in the long one.
Western Asia/SW Asia, NE & Eastern Africa
1987 - 2017
25,000 - 100,000
1988 - 2015
Stable/ Fluctuating
CSR7
References
Perennou, C.P., Mundkur, T. and Scott, D.A. 1994. The Asian Waterfowl Census 1987-1991: distribution and status of Asian waterfowl. IWRB Spec. Publ. No. 24; AWB Spec. Publ. No. 86. Slimbridge, UK and Kuala Lumpur, Malaysia.
Sheldon, R. 2017. Estimates of breeding waterbird populations in Central/SW Asia, The Caucasus and the Arabian Peninsula.
Wetlands International (2017) Flyway trend analyses based on data from the African-Eurasian Waterbird Census from the period of 1967-2015. Ede, The Netherlands: Wetlands International. URL: http://iwc.wetlands.org/index.php/aewatrends
Notes
Stable/fluctuating in the long-term. This overall trend confounds large long-term fluctuation.
West Indies
2006 - 2006
30 - 60
1991 - 2001
Increasing
WPE5
References
Peterjohn, Bruce, in litt., April 2002.
Unpublished information supplied by Wetlands International Specialist Groups, 2006.
Notes
Population added in WPE3.
Colonised in early 1990s, (529). 10-20 pairs (30-60 individuals). See 336.
South Asia
—
100,000 - 200,000
—
Unknown
WPE5
References
Asian Waterbird Census unpublished data. 2002.
Notes
20: 48,000 counted in 2002.
E, SE Asia
2001 - 2001
100,000 - 1,000,000
1981 - 1991
Stable
WPE5
References
Chan, Simba, In litt,. 2002. Unpublished information on Asian waterbirds.
nigripes
1987 - 1991
25,000 - 1,000,000
—
Unknown
WPE5
References
Perennou, C.P., Mundkur, T. and Scott, D.A. 1994. The Asian Waterfowl Census 1987-1991: distribution and status of Asian waterfowl. IWRB Spec. Publ. No. 24; AWB Spec. Publ. No. 86. Slimbridge, UK and Kuala Lumpur, Malaysia.
immaculata
2002 - 2004
25,000 - 100,000
—
Unknown
WPE5
References
Bishop, K.D. 2003. A Review of the Avifauna of the TransFly Eco-region: the status, distribution, habitats and conservation of the region?s birds. Report to WWF South Pacific Program.
Jaensch, Roger, August 2005. Detailed data and notes on numbers and distribution of waterbirds in Australia, updated from Jaensch, 2003, and derived from more than 90 Australian sources including Wetlands International, Birds Australia and Jaensch, 2003,
Notes
Included in nigripes by Marchant & Higgins (1990).
Estimate based on review of previous information and some recent information, which suggest that total numbers in Australia and S New Guinea are between 25,000 and 100,000, assuming most of the S New Guinea birds are immaculata. If immaculata and nigripes are combined as one population, estimate may be better expressed as a range of 25,000-1,000,000 in view of WPE4 data for nigripes.
Egretta gularis
Western Reef-egret
gularis
West Africa
1991 - 2014
10,000 - 50,000
1997 - 2014
Stable/ Increasing?
CSR7
References
Dodman, T. 2014. Status, Estimates and Trends of Waterbird Populations in Africa: AEWA-listed African populations. Wetlands International. (CSR6 African populations) URL: https://www.wetlands.org/publications/1304/
van Roomen M., Nagy S., Foppen R., Dodman T., Citegetse G. & Ndiaye A. 2015. Status of coastal waterbird populations in the East Atlantic Flyway. With special attention to flyway populations making use of the Wadden Sea. Programme Rich Wadden Sea, Leeuwarden, The Netherlands, Sovon, Nijmegen, The Netherlands, Wetlands International, Wageningen, The Netherlands, BirdLife International, Cambridge, United Kingdom &, Common Wadden Sea Secretariat, Wilhelmshaven, Germany. URL: http://www.waddensea-secretariat.org/sites/default/files/downloads/status_coastal_birds_eaf_2014_1.pdf
Wetlands International (2017) Flyway trend analyses based on data from the African-Eurasian Waterbird Census from the period of 1967-2015. Ede, The Netherlands: Wetlands International. URL: http://iwc.wetlands.org/index.php/aewatrends
Notes
This form and schistacea sometimes treated as separate species, Western Reef Heron.Sometimes assigned to Egretta garzetta.
Review of more recent data, including 2013 and 2014 counts
Van Roomen et al (2015) found increasing trend based on the IWC data. Wetlands International (2017) found that the long-term trend is stable/fluctuating, the short-term is uncertain. Wetlands International's assessment agrees well with Dodman (2014).
schistacea
North-east Africa & Red Sea
1937 - 2011
10,000 - 15,000
2014 - 2014
Declining/ Stable
CSR7
References
Dodman, T. 2014. Status, Estimates and Trends of Waterbird Populations in Africa: AEWA-listed African populations. Wetlands International. (CSR6 African populations) URL: https://www.wetlands.org/publications/1304/
Nagy, S., Alanazi, F., Almomen, A. Alsuhaibani, A, AlRashidi, M., Dereliev, S., Keijl, G. Ruiters, P. & Shobrak, M. 2014. Winter waterbird survey in the Kingdom of Saudi Arabia in January 2014. Wetlands International, Ede, The Netherlands.
Sheldon, R. 2017. Estimates of breeding waterbird populations in Central/SW Asia, The Caucasus and the Arabian Peninsula.
Wetlands International (2017) Flyway trend analyses based on data from the African-Eurasian Waterbird Census from the period of 1967-2015. Ede, The Netherlands: Wetlands International. URL: http://iwc.wetlands.org/index.php/aewatrends
Notes
Sometimes assigned to Egretta garzetta schistacea.
See CSR6 and Sheldon (2017).
Dodman (2014) assumed that the population is stable in the absence of human impacts along the Red Sea coast. Reviewing of available IWC data and the formal trend analysis suggest that a steep decline might have taken place between 1990 and 2015 (Wetlands International 2017). This is probably driven by destruction of coastal wetlands and mangroves particularly along the northern coast of the Red Sea (Nagy et al. 2014).
South-west Asia & South Asia
1990 - 2012
10,000 - 25,000
2006 - 2015
Stable?
CSR7
References
Sheldon, R. 2017. Estimates of breeding waterbird populations in Central/SW Asia, The Caucasus and the Arabian Peninsula.
Wetlands International (2017) Flyway trend analyses based on data from the African-Eurasian Waterbird Census from the period of 1967-2015. Ede, The Netherlands: Wetlands International. URL: http://iwc.wetlands.org/index.php/aewatrends
Notes
Sometimes assigned to Egretta garzetta schistacea. Sometimes assigned to asha.
See CSR6 and Sheldon (2017).
The short-term trend is uncertain but apparently stable.The long-term one is strong increase.
dimorpha
Madagascar
—
6,000 - 20,000
—
Declining
WPE5
References
Hawkins, F. In litt. 2002. Unpublished notes on waterbird population estimates of Madagascar.
Aldabra & Amirante Is
—
3,000 - 9,000
1991 - 2001
Stable
WPE5
References
Dodman, T. 2002. Waterbird Population Estimates in Africa. Unpublished report to Wetlands International.
Rocamora, G. and Skerrett, A. 2001. Seychelles. Pp 751-768. In L.D.C. Fishpool and M.I. Evans, eds. Important Bird Areas in Africa and associated islands: Priority sites for conservation. Newbury and Cambridge, UK: Pisces Publications and BirdLife International
Coastal Eastern Africa
1996 - 1996
15,000 - 20,000
1991 - 2001
Stable?
CSR7
References
Baker, N. (compiler). 1996. Tanzania Waterfowl Count, January 1995: The first coordinated count of the major wetlands of Tanzania. Wildlife Conservation Society of Tanzania. Dar es Salaam
Dodman, T. 2014. Status, Estimates and Trends of Waterbird Populations in Africa: AEWA-listed African populations. Wetlands International. (CSR6 African populations) URL: https://www.wetlands.org/publications/1304/
Notes
An earlier figure of 10,000 was erroneously used based on the same reference.
No monitoring data is available. Trend assessment is based on circumstantial evidence.
Egretta sacra
Pacific Reef-egret
sacra
2006 - 2006
100,000 - 1,000,000
1985 - 2005
Stable?
WPE5
References
Unpublished information supplied by Wetlands International Specialist Groups, 2006.
Notes
Regarded as 'rapidly declining and very rare' in China (125).
Regarded as 'rapidly declining and very rare' in China (125).
albolineata
—
1,000 - 2,000
1990 - 2000
Stable
WPE5
References
Barr?, N. and Dutson, G. 2000. Oiseaux de Nouvelle-Caledonie. Liste commentee. Suppl. Alauda (68), 3: 49pp.
Notes
Population added in WPE3.
Egretta eulophotes
Chinese Egret
E, SE Asia
2000 - 2010
3,000 - 4,100
—
Stable
WPE5
References
BirdLife International 2001. Threatened Bird of Asia: the BirdLife International Red Data Book. Cambridge, UK. BirdLife International
Zhiying, D. 2009. The Chinese Egret, Feathers of snow in the summer sky. Chinese National Geography Vol. 1, Issue 2. Bulletin of Academy of Science Pyeongyang.
Notes
Hwang (2011) estimates 1,000-1,600 breeding individuals in South Korea from 2000 to 2010, 1,000-1,200 (Collar 2001) or 1,200-1,500 (BASP 2004) in DPRK, and 1,000 birds in China (Zhiying 2009).
The Heron Red List (Version 2024-2) is shown below.
This authority for the global classification of the conservation status of species is the IUCN. It is published periodically as the IUCN Red List of Threatened Species. BirdLife International is the listing authority for birds. HeronConservation partners with BirdLife International on its periodic assessments for herons. The Heron Red List is the same as the IUCN list. Like all species, herons may be classified the following categories, LC = Least Concern, NT = Near Threatened, VU = Vulnerable, EN = Endangered, CR = Critically Endangered, EW = Extinct in the Wild, and EX =Extinct. More information on the meaning of these categories may be found in the IUCN Red List Categories & Criteria document.
The IUCN process is conducted at the species level, whereas HeronConservation does its assessments at a population level. So for some species the classification of a species may differ from that of one or more of its populations. For information on the listing assessment process, please see the IUCN Website.
The IUCN Assessment in some cases identifies species, uses a listing order, and uses scientific and English names that are at times deviant from those used by HeronConservation. There are a number of reasons for these differences, mainly that HeronConservation, being concerned only with one group of species can tracks and implement changes within its group more readily, whereas IUCN needs to maintain a more conservative approach to such changes across its periodic assessments and needs to maintain a common global process. As there is not a problem in identifying which species is being referred to, HeronConservation has chosen to retain the IUCN order and names to maintain its consistency with the Global Red List.
HeronConservation will continue to partner with BirdLife in its evaluation of the status of heron species. You are welcome to participate in this process by communicating your data and information to the HeronConservation Co-chairs or Steering Committee Members.
1 New Jersey Audubon, Bernardsville, NJ 07924, USA; 2 Lenoir-Rhyne University, Hickory, NC 28601, USA; 3 New York City Audubon Society, New York, NY 10010, USA; 4 North Carolina State University, Raleigh, NC 27695, USA; 5 Friends University, Wichita, KS 67213, USA; 6 North Carolina Wildlife Resources Commission,Raleigh, NC 27606, USA
State-of-the-art satellite GPS transmitters (48-g Bird Solar; e-obs) are providingunprecedented details regarding movements and habitat utilization for birds heavier than 1 kg. We present current information on habitat utilization and the spatial distributions of 13 Great Egrets (Ardea alba) captured in various locations from 2013 to 2015. Nearly 560,000 GPS points were used to quantify the size of areas used by birds during 3694 observation-days by using LoCoH program in the R platform. The habitat size required for breeding (n = 10 birds), particularly when young require most food, islarger than that used for the post-breeding period (n = 7) and during winter (n = 5). All major migrations occurred at night, and we present details for post-breeding (n = 9) and Spring (n = 5) migration segments. Two young-of-the-year, non-breeding birdsdisplayed both conservative and exploratory use of habitat.
Estimating the breeding population of Great Blue Herons in Maine: what's not on our colony list?
D'Auria, Danielle E.1; Otto, Mark C.2
1 Maine Department of Inland Fisheries and Wildlife, Bangor, ME 04401, USA; 2 U.S. Fish and Wildlife Service, Patuxent Research Refuge, Laurel, MD 20708, USA
The Great Blue Heron (Ardea herodias) is listed in Maine as a Species of Special Concern due to a decline in nesting pairs and colonies along the coast since the mid-1980s. In order to determine whether the decline is limited to the coast, occurring statewide, or due to movement from the coast to inland sites, an aerial survey was conducted in 2015 to obtain an estimate for the statewide breeding population. Aerial surveys had been conducted previously, but this time a stratified dual-frame design accounting for imperfect detection of colonies was used. The strata were based on habitat and known colony densities. The area frame consisted of 10 km x 10 km plots; a sample of which was searched independently by front and rear observers for both new and known colonies. The list frame consisted of plots that contained known colonies and was only searched for known colonies. Detection of active nests within colonies and colonies themselves were accounted for in the estimation. The estimates of total nests and active nests were obtained from the number of colonies multiplied by the average colony size. The estimate of total colonies for the state is 336 (range = 261-455 colonies) and the estimate of active nests is 1,800 (range = 1,631-2,159 nests). Observer detection ranged from 31 to 95 percent. The list coverage, or the percentage of colonies included in the list frame, was 67 percent but varied widely among strata. By repeating these methods at future intervals (e.g., every 5 years), we plan to obtain population trends for each strata as well as the entire state of Maine.
Conservation status of herons in Paraguay: future scenarios in face of accelerated habitat degradation
del Castillo, Hugo1; Clay, Rob2; Lesterhuis, Arne1; Yanosky, Alberto1
1 Guyra Paraguay, Parque Ecológico del Rio, BioCentro, Avda. Carlos Bóveda s/n., CC 1132, Viñas Cue, Asunción, Paraguay; 2 Director of the Western Hemisphere Shorebird Reserve Network (WHSRN)
Paraguay’s five major ecoregions hold a total of 14 heron species. Currently, no species is considered threatened at a national or international level; in fact on the contrary, most species are common to abundant. Exceptions include the Boat-billed Heron, which is scarce and locally distributed, but probably overlooked due to its nocturnal habits; andthe three species of bittern (Pinnated, Least and Stripe-backed). Knowledge of the vocalizations of the two Ixobrychus bitterns has revealed them to be more widespread and abundant than previously thought, though they are still infrequently recorded and primarily found in wetlands within the Paraguay River valley. A recent increase in Pinnated Bittern records is likely related to the expansion of rice fields, and the relative ease of observing the species in such habitat. Capped Heron is the one species with a restricted distribution, limited to the Pantanal and associated wetlands in the north of the country. Little Blue Heron is a scarce vagrant to the country, though records appear to be increasing. Cattle Egret was already widespread in Paraguay by 1977, when first recorded by ornithologists. There is increasing evidence for seasonal movements by both that species and Striated Heron. While most herons remain common and widespread throughout the country, the increasing loss of wetland habitats, and their degradation through runoff and sedimentation (driven by massive deforestation and conversion to industrial agriculture) is of concern. Although the rapid and ongoing expansion of rice agriculture in Paraguay provides important foraging habitat for at least 8 heron species, their populations depend on the survival of natural wetland and woodland habitats in surrounding areas for roosting and breeding.
Development of a survey protocol for monitoring Reddish Egrets in Florida
Cox, Andrew; Schwarzer, Amy
Fish and Wildlife Research Institute, Florida Fish and Wildlife Conservation Commission, Gainesville, FL, 32601, USA
The Reddish Egret (Egretta rufescens) is North America’s rarest heron and roughly 10% of its global population resides in Florida. The rarity of the species, its localized breeding distribution and a potential negative population trend have prompted the need to monitor thisspecies. However, standard aerial surveys performed from fixed-wing aircraft often implemented for colonial nesting wading birds are ineffectual for this dark-plumaged and sub-canopy nesting species, leading some researchers to use either direct counts or flight-line surveys. During the 2015 breeding season we compared the efficacy of flight-line surveys and direct counts using data collected by multiple observers during repeated visits to colonies located in three of Florida’s core breeding areas (Tampa Bay, Florida Bay, and the lower Florida Keys). Our objectives were to 1) determine the appropriate duration of flight-line surveys, 2) estimate variation between multiple flight-line surveys performed at the same sites, and 3) estimate the correlation between flight-line surveys and direct counts. Average variation between counts was high for a 1-hr flight line survey (60%), but more acceptable for 2-hr (20%) or 3-hr (16%) surveys. Flight-line surveys often produced abundance estimates that were biased high when compared to direct counts, likely because nest exchanges occurred >1 time per survey, violating a primary assumption of the method, or because breeding sites contained interior ponds used for foraging by adult birds breeding elsewhere. Nevertheless, some breeding sites were not amenable to direct counts and others are in areas where such access is prohibited. Ultimately, variation in habitat, site access, and breeding synchrony will necessitate the use of multiple approaches when conducting a statewide survey.
Range-wide Survey of Inland Wading Bird Nest Colonies in North Carolina
Cook, K. L.1; Schweitzer S. H.1; Anderson S.1; Cameron, S. E.2; Shaiyen, M.1
1 N.C. Wildlife Resources Commission, Raleigh, NC 27606, USA; 2 U.S. Fish & Wildlife Service, Asheville, NC 28801, USA
Although fairly common in North Carolina, accurate estimates of Great Blue Heron (Ardea herodia) andGreat Egret (A. alba) abundance are poor. Since the 1970s, wading bird colony surveys have been conducted only for certain regions. From April to mid-May, 2008-2012, we completed a stratified aerial survey of all major waterways within the state breeding range. Goals were to design a repeatable survey accounting for detection probability using a double-observer method. We also wished to evaluate landscape characteristics associated with persistent and abandoned heronries to improve conservation recommendations. A total of 490 active heronries were detected ([192] Piedmont, [298] Coastal Plain), 80% of which had not been recorded previously. Heronry size was not correlated with detection probability and detection probability did not differ between observers (0.9, 95% credible interval [CI] [0.8-1.0]). Along our flight route, the Bayes estimate of total heronry abundance incorporating detection probability was 548 heronries (95% CI [535–561]). We estimated 533 heronries that included GBHE (95% CI: [520–546]) and 38 heronries with GREG (95% CI: [37–39]). Nest abundance estimates were 8,306 GBHE nests (95% CI [7,963–8,660]) and 4,963 GREG nests (95% CI [4,963–5,406]). Other species had low counts and or detectability and so are not reported.
Hydrologic fluctuations influence dailysurvival rates of small herons and Great Egrets in a subtropical littoral wetland
Essian, David A.1; Chastant, Jennifer1; May, Jenna2; Gawlik, Dale E.2
Subtropical freshwater wetlands in South Florida USA, support large populations of wading birds that are thought to be food-limited. The production of many wading bird prey species is controlled by hydrological variation, which is increasingly regulated by human activities through water management regimes. Thus, we expected that water management regimes on Lake Okeechobee, Florida USA would also influence wading bird reproduction. We use a model selection approach to investigate the influence of hydrological parameters on the daily nest survival rates (DSR) of wading birds. We pooled Tricolored Heron (Egretta tricolor) and Snowy Egret (Egretta thula) nests since they are indistinguishable during the incubation period. Because lake stage and water level recession rates, in particular, can be adjusted by water managers, we focused on the influence of those parameters on the DSR of Great Egrets (Ardea alba; n=298) and small herons (n=1,524). Nest survival for all species was highest when lake stage was below 4.0 m and above 3.6 m. Lake stage was the only important parameter that influenced DSR of Great Egrets, but this model was only marginally better than the null model. Water level recession rate, lake stage, and their interaction were important parameters for predicting DSR of small herons, regardless of nesting stage. These species were more sensitive to lake stage and water level recession rate than were Great Egrets. Our results suggest that water management strategies could be designed to benefit species sensitive to hydrologic constraints such as small herons without hindering the reproductive success of less sensitive species.
Current Status, ecological characteristics and conservation of Family Ardeidae in natural and disturbed forests of urban area, Korea
1 Urban Planning Research Group, Daejeon Development Institute, 34863 Daejeon, Korea; 2 Korea Institute of Environmental Ecology Inc., 34014 Techno 1-ro, Yusunggu, Dajeon, Korea; 3 National Institute of Ecology, 33657,Geumgang-ro, Maseo-myeon, Seocheon-gun, Chungcheongnam-do, Korea; 4 Department of Sustainable Development, Yeungnam University, 712-749, Gyeongsan, Korea
Of 72 egret species recorded in the world, 18 species were observed in Korea. According to the nationwide census conducted by the National Institute of Environmental Research between 2011 and 2012, 35,512 breeding pairs at the 148 sites were recorded in Korea. We found 142 breeding pairs of 5 species at 3 sites in Daejeon in 2016. 10 individuals (5 gray herons, 4 great egret and 1 intermediate egret) were tagged with GPS-Mobile based Telemetry (WT 300; 35 x 63 x 14 mm, >27 g, Korea) and released. Some of them (5 gray herons, 2 great egret) were successfully tracked by now. Their home-range sizes were about 20-25 km2 (100% Minimum Convex Polygon Method, 95% Kernel Density Estimation) in breeding season. They moved down to the southern part of China and Vietnam on October 2015. As a result of analyzing potential habitats using 12 variables selected by Maxent model, 106.68 km2 (19.7%) were extracted in Daejeon, Korea. Among them, we selected Wolpyung Park as compensatory habitat, and placed 20 decoys with nests anda recorder stored breeding song on the top of trees to induce egret breeding on January 2016, but failed. More effective long-term plan should be needed to solve the conflict between residents and breeding egrets.
Dry rice paddies cause a decline of the breeding herons and egrets in Italy
1 Dipartimento Scienze Terra Ambiente, Pavia, Italy; 2 IGG-CNR, Pisa, Italy; 3 IREA-CNR, Milano, Italy
After a period of strong increase since 1980, and a peak around 2000, the breeding herons and egrets in Northwestern Italy entered a phase of decline. The population fluctuations were affected by several climatic factors, but the main cause for the increase was reduced human-induced mortality. The recent decline, however, occurred following the expanding practice of rice cultivation on dry paddies that during 2015 reached 80% of the rice surface in some areas. The dry paddies are not flooded for more than a few days, and become unsuitablefor the herons and egrets, except for the Cattle Egret, the only one of the seven species that is adapted to forage on dry lands and that is still increasing in our study area. In the paddies that remain flooded, prey availability for herons has changed dramatically due to arrival of alien species and to the decreased water level compared to the traditional practices. In the other European areas of rice cultivation, in Spain, France, and Greece, cultivation without submersion has not been adopted yet, but in Italy the new practices are undermining the value of rice cultivation for waterbirds. The monitoring of the 200 heronries in our study area, now in its 45th year, is continued thanks to a group of 100+ volunteer collaborators.
Representing hydrologic variability in heron models: key processes for wetland ecosystem management
The response of herons and other wading birds to hydrologic variability has been studied for over 50 years in the Florida Everglades (USA) and globally. Collectively, these studies illustrate a variety of mechanisms through which hydrologic variability affects nesting and foraging, and how this variability can be represented in ecological models that have application to wetland restoration and management. In wetlands with a pronounced seasonal water cycle, herons and other wading birds often initiate nesting when water levels are dropping and aquatic animals are concentrated into shallow water. The success of nesting is dependent on the rate of receding water being at an optimal level, with rates above andbelow the optimum causing lower nest survival. When water levels stop receding and rise suddenly due to rain or inflows, birds will abandon their nests. This pattern contrasts with that of floodplain wetlands where wading birds often initiative nesting when water levels are rising and prey are dispersing rather than being concentrated. Juvenile aquatic animals disperse out of river channels to the floodplain where they experience a rapid growth rate leading to an increase in prey biomass for birds. The parameters hydroperiod and minimum water level affect the production of prey populations, establishing the upper limit to subsequent foraging conditions, with the actual value being lowered by the degree to which deep water levels restrict access to prey by birds. The range in water level fluctuations defines the spatial extent of habitat that becomes suitable for foraging. In floodplain systems this variable is often the primary determinant of numbers of nests.
Characterization of Ardeid assemblages on the southern coast of Cuba
1 Facultad de Biología, Universidad de La Habana; 2 Centro Nacional de Áreas Protegidas; 3 Empresa para la Protección de la Flora y Fauna, Reserva Ecológica “Los Pretiles”; 4 Centro de Investigación y Servicios Ambientales, ECOVIDA; 5 Museo de Historia Natural “Tranquilino Zandalio de Noda”; 6 Empresa para la Protección de la Flora y Fauna, Refugiode Fauna “Punta Caribe”; 7 Instituto Nacional de Ciencias Agrícolas “Los Palacios”; 8 Empresa para la Protección de la Flora y Fauna, Refugio de Fauna “Canales del Hanábana”; 9 Empresa para la Protección de la Flora y Fauna, Refugio de Fauna “Tunas de Zaza”; 10 Empresa para la Protección de la Flora y Fauna, Refugio de Fauna “Delta del Cauto, Granma”; 11 Empresa para la Protección de la Flora y Fauna, Refugio de Fauna “Delta de Cauto, Las Tunas”, Cuba
Cuba has 12 species of herons distributed throughout the national territory, with exception of American Bittern, all breeding in Cuba and has migrant populations from North America. The group is well represented in natural and anthropic wetlands, however little has been published about their status and distribution. In this work weevaluated the ardeidos assembly in nine Cuban natural wetlands distributed along the southerncoast of Cuba from May 2011 to March 2013,and one ricefield between 1992 and 1995. Of the 12 species represented in the archipelago, nine were presentin 100% of observations in nine of the wetlands studied. Here were included all representatives of genusEgrettain Cuba. The highest values of abundance were recorded in Los Palacios (1,188 individuals); Delta del Cauto (958 individuals) and Canales Hanábana (924 individuals). In more than 50% of the sites Snowy Egret, Little Blue Heron, Tricolored Heron and Great Egret were identified as the most abundant species. In Punta Caribe and Tunas de Zaza, the highest values of abundance were recorded in the period of Permanent Residence (May-June), while in Los Palacios y Monte Cabaniguán was during the fall migration (October-November). In the remaining sites these higher abundance values were reported during the spring migration (February-March). Meanwhile, in the ricefield 12 species of herons were identified. Of these, the most abundant were Cattle Egret, Snowy Egret, Little Blue Heron and Great Egret. In general,the results support the importance of natural and anthrogenic wetlands in Cuba for ardeidos.
Clinal variations in dark morph proportion of Reddish Egret (Egretta rufescens) in Cuba
1 Departamento Biología Animal y Humana, Facultad de Biología, Universidad de La Habana; 2 Museo de Historia Natural Felipe Poey, Facultad de Biología, Universidad de La Habana, Cuba
The Reddish Egret presents a polymorphic plumage that is independent of age and sex. According to information from the ratio of dark morph in some areas of its range, the species appears to exhibit a cline variation, with dark individuals predominating in the most western populations (e.g., Baja California, ~100% dark morph), while the eastern populations contain a small proportion of these (e.g., Great Inagua, ~12% dark morph). An analysis of the proportion of dark individuals over 20 Cuban wetlands revealed a consistent pattern of geographic variation with apparent cline described in the global distribution of the species. In turn, the proportion of dark individuals was higher in the N coast than in the S, but in both cases the probability of observing a dark individual decreased with distance from the western end of Cuba (cline west-east, coast N: 92 50%, S coast: 80-9%). A case study in three wetlands of the south coast distributed throughout Cuba confirm the results found for this coast. The study included systematically sampled in these wetlands for two years.
A decade of Reddish Egret research: looking back and moving forward
Green, M. Clay1; Ballard, Bart M.2
1 Department of Biology, Texas State University, San Marcos, Texas 78666, USA; 2 Caesar Kleberg Wildlife Research Institute, Texas A&M University-Kingsville, Kingsville, Texas 78363, USA
In 2006, the US Fish and Wildlife Service published a species status report for the Reddish Egret (Egretta rufescens), the first status review of the species in 15 years. From that report, it was evident that knowledge gaps on the ecology and limitingfactors of the species remained. Since 2006, we have conducted research in Texas and various parts of the species’ range on 1) movement ecology, 2) nesting and foraging ecology, 3) juvenile and adult survival and 4) genetic differentiation and gene flow. Juvenile Reddish Egrets exhibit nomadic behavior during post-fledging dispersal before showing little movement during first winter and subsequent breeding season, whereas roughly 60% of adult Reddish Egrets nesting in Texas exhibit migratory behavior overwintering in Mexico and as far south as El Salvador. While annual survival of adult birds is high, survival of juvenile Reddish Egrets is low (~25%) and potentially as low as 10% during first 3-6 months post hatch. Low juvenile survival is in contrast to high nesting survival (~80-90%) from incubation through 4-5 weeks post hatch. Within Texas, we found little genetic differentiation (mtDNA and microsatellites) geographically or between color morphs. Across the range, we found significant differentiation along a longitudinal gradient with genetically isolated population centers in Baja California, Chiapas and Bahamas. Populations in Louisiana, Texas, eastern Mexico and Florida appear to be transitional between the extremes of Baja and Bahamas. Our research fills many information gaps for this little studied species, provides important implications for future research, and informs resource managers for more directed and impactful conservation for the Reddish Egret.
HeronryMAP:Africa — mapping the distribution and status of ardeid (and other waterbird) breeding colonies in Africa
Harebottle, Doug M.
School of Natural and Applied Sciences, Sol Plaatje University, Kimberley, South Africa
Colonial breeding waterbirds are spread across seven bird families — Laridae, Phalacrocoracidae, Ardeidae, Phoenicopteridae, Threskiornithidae, Pelecanidae and Ciconidae. Due to their conspicuous behavior and often socio-economic and ecological impacts, most taxa have been well studied. In Africa, information on the status and distribution of breeding colonies in the Ardeidae is severely lacking which provides a gap in the knowledge of how important these sites are in terms of location and productivity. HeronryMAP: Africa is a citizen science initiative that aims to address this gap through the systematic collection of long-term dataon inter alia where ardeid breeding sites occur, their species composition, nest abundance and site tenureship. Preliminary results are presented from 2013, and challenges identified and discussed regarding data mobilization and sustainability. Future objectives such as the assessment of priority sites, and identification of conservation action for colonies under threat are discussed and the impact of climate and landscape changes are briefly highlighted.
Rice fields support the largest known breeding population of the endangered Australasian Bittern
Herring, Matthew W.1,2; Zander, Kerstin K.2; Garnett, Stephen T.2
1 Bitterns in Rice Project, Leeton, NSW 2705, Australia; 2 Charles Darwin University, Darwin, NT 0909, Australia
The Australasian Bittern (Botaurus poiciloptilus) is a poorly known, globally endangered species with a total population of just 1,000-2,499 mature individuals. The Riverina region of New South Wales is recognised as a stronghold. It supports around 95% of Australia’s rice production, which constitutes approximately 100,000 hectares in years of 100% water allocation. Despite this, little was known about the Australasian Bittern population found in these rice fields. From 2012-2016, standardised surveys of rice crops on randomly selected farms in two of the three main rice-growing areas wereused to estimate the population size and determine the extent of breeding. Occupancy at the 23-30 hectare sites ranged 0.23-0.29, depending on the year and region. Most observations were of one or two birds, but up to four were recorded during a single survey. Habitat occupancy modelling, accounting for the unsurveyed third region and substantial detectability issues, suggests that in most years these rice fields attract approximately 500-1,000 mature individuals. This represents around 43% of the estimatedglobal population. Nests found at the randomly selected farms indicate widespread breeding and observations of fledged young confirm that some nests succeed before harvest. The results highlight the overlooked conservation role of agricultural wetlands inAustralia and the potential for dual-purpose water use. Bitterns showed a strong preference for more traditional rice growing methods of aerial sowing with early inundation, as opposed to direct-drill sowing with delayed inundation. However, driven by water savings, an increasing number of rice growers are altering their sowing methods and water management. Development of bittern friendly rice growing incentives will be discussed.
Impacts of a road construction on water bird populations and first regional rehabilitation actions at Asunción Bay Ecological Reserve (Paraguay)
Guyra Paraguay, Parque Ecológico del Rio, BioCentro, Avda. Carlos Bóveda s/n., CC 1132, Viñas Cue, Asunción, Paraguay
The Asunción Bay, is a relatively small bay (c.600 ha in total) located along the northern outskirts of Asunción, the capital of Paraguay. Although a relatively small area, more than 290 species of birds have been recorded in the bay, including 89 waterbird. Families represented by most species include Scolopacidae (18 species), Anatidae (15 species), Rallidae (13 species) and Ardeidae (11 species). 34 of the recorded waterbirds are migratory species, including 21 Nearctic migrants and 13 Austral migrants. Due to its importance for migrants in particular, the bay has been designated as an IBA and WHSRN site, and in 2005 was declared as an Ecological Reserve. The recent development of a coastal road has presented important opportunities for the urban population of Asunción to reconnect with its natural heritage, but unfortunately dredging to create the embankment for this road in 2010 destroyed about 70% of the habitat in the bay. The disappearance of muddy beaches on the bay caused drastic reductions of the total number of Nearctic shorebirds that regularly visited the Bay, but also caused changes in abundance and diversity of resident waterbird populations. The Municipality of Asunción and the Ministry of Public Works have been working together with Guyra Paraguay on the implementation of a series of habitat recuperation and management measure to improve remnants habitats for both waterbirds in general and shorebirds in particular.
Intraspecific and Intersexual Variation in Three Species of Wading Birds
Maccarone, Alan D.1; Brzorad, John N.2
1 Biology Department, Friends University, Wichita, Kansas, USA; 2 Reese Institute for Conservation of Natural Resources, Lenoir-Rhyne University, Hickory, North Carolina, USA
Intraspecific and intersexual morphological variation is common in many groups of birds, but few data regarding such differences exist for Ardeids. Since 2008, we have trapped long-legged wading birds in Kansas and along the East Coast for telemetry studies. Captured individuals are weighed and several measurements taken before they are released. Beginning in 2013, a blood sample was collected from each bird, which was used to determine its sex. Measurements of 103 birds of three species were used to examine relationships among mass, culmen length, and tarsus length, and to determine whether males of each species differ from females in these values. Great Blue Herons (1,702-2,859 g), Great Egrets (1,769-1,300 g), and Snowy Egrets (349-539 g) all showed high variation both in body weights and morphometric measurements. For all three species, weight was correlated significantly both with culmen and tarsus lengths, which were themselves strongly associated. For Great Egrets and Snowy Egrets, males were significantly heavier than females and also had longer culmen and tarsus lengths (all P <0.003). Sample size for Great Blue Herons was not large enough to compare male and female birds. We discuss the possible implications of intraspecific variation and sexual differences in Ardeids.
Foraging Microhabitat Selection by Long-legged Wading Birds at an Artificial Weir
Maccarone, Alan D.; Renken, Rachel C.; Thompson, Jeane A.
Biology Department, Friends University, Wichita, Kansas, USA
To better understand how wading birds select among different foraging microhabitats that show spatial heterogeneity, we divided a 100-m-long concrete weir located at the terminus of the Little Arkansas River in Wichita into 10 patches based on water depth. We observed four species of wading birds during 60 1-h periods from 12 June-28 July 2015 to document microhabitat use and feeding behavior. The independent variables collected before each session were time of day, date, water level, water clarity, and flow velocity. We documented capture efficiency, prey length (relative to bill length), and aggressive interactions for Black-crowned Night-Herons (n = 396), Great Egrets (n = 54), Snowy Egrets (n = 36), and Great Blue Herons (n = 30). Time of day and water level were the only significant predictors of weir attendance. A total 348 fish were captured, of which 108 were 3/4 bill length or greater. Wading bird species differed in capture efficiency, mean prey lengths, and the primary patch use. Great Blue Herons and Black-crowned Night-Herons captured mainly large fish (gizzard shad, catfish, and freshwater drum); Great Egrets captured both large and small fish, and Snowy Egrets captured mainly small fish (minnows). Overall aggression rate was correlated with the number of large fish captured but not with total fish, whereas the per capita aggression rate was correlated with the number of birds at the weir. Electro-fishing showed a non-uniform distribution of fish among the 10 patches. The pattern of fish spatial distribution was reflected in prey-capture patterns, where mean fish length differed significantly by patch and ranged from 2.8 cm to 11.9 cm.
Recent trends of the nesting location of Grey Herons in Hokkaido, northern Japan
Matsunaga, Katsutoshi
Hokkaido Grey Heron Research Group, 5-4-8-103, Hachiken 8-jo Higashi, Nishi-ku, Sapporo, Hokkaido 063-0868, Japan
The nesting location of Grey Herons (Ardea cinerea) in Hokkaido (83,450 km2), northern Japan was documented from 1960 to 2016. During the study period, 152 colonies were confirmed to be exist, though some of them have been already abandoned. All herons were observed to nest at trees in the inland area until the early 1990s. After the mid-1990s, however, 10 colonies were confirmed in unusual locations: an islet, flooded trees and buoys in reservoirs, and offshore rocks. These type of locations could be considered to be selected to defend their eggs and chicks against terrestrial predators by being surrounded with water area. In fact, in Hokkaido, Brown Bears (Ursus arctos) and Common Raccoons (Procyon lotor) were observed to eat chicks in conventional colonies in 1994 and 2012, respectively. Moreover, some colonies were recently established in isolated groves in towns and others adjacent to houses despite the presence of vast woodlands around. It would be another strategy to prevent bears and raccoons from approaching the colony. The number of colonies had increased, at least until 1999, and raccoons, which are alien species, have rapidly expanded their range until now. These situations would increase the encounters between herons and predators, and therefore, herons may be forced to nest in unusual locations.
The effects of colony structure and nest position on the reproductive success of small herons
May, Jenna C.; Essian, David A.; Gawlik, Dale
E. Florida Atlantic University, Boca Raton, FL 33431, USA
When food is not limiting, competition for high quality nesting sites can limit the density of breeding birds. Quality nesting sites must provide structural support for nests as well as to offer effective protection against predators and unfavorable weather. Anthropogenic influences can alter the structure and composition of vegetation available for nest sites, thereby providing birds with novel habitat. During the 2015 breeding season, we examined the effects of colony structure and nest position on the reproductive success of Tricolored Herons (Egretta tricolor) and Snowy Egrets (Egretta thula) nesting at Lake Okeechobee, Florida USA, to determine if colonies in anthropogenic habitat (spoil islands) had lower reproductive rates than colonies in natural habitat (willow, Salix sp.; islands). Daily survival rate (DSR) of nests did not differ significantly between the two colony types. The best model predicting DSR included nest height, distance to canopy, and type of substrate species (invasive or noninvasive). Apparent survival was higher for nests placed farther from the canopy edge, higher from the ground, and in noninvasive substrate. Results indicate that spoil islands are capable of providing nesting habitat comparable to natural islands in some years. However, factors that led to a lower DSR, (e.g., invasive plants and short vegetation), also tended to be more prevalent on spoil islands, so over a longer time we would expect to see differences in DSR emerge.
Diversity of waterbirds in Periyakulam Lake, Tiruchirappalli District, Tamil Nadu, Southern India
Mohanraj, Sivanantham; Pandiyan, Jeganathan
PG Research Department of Zoology and Wildlife Biology, AVC College (Autonomous), Mannampandal – 609 305, Mayiladuthuari, Tamil Nadu, India
The bird community of Periyakulam wetlands in Tiruchirapalli District, Tamil Nadu, Southern India was studied during January 2011 to December 2012. The methodology followed was mainly observations using binoculars, andthe site was done by direct count. A total of 35 species belongingto 7 orders and 18 families, including 14 resident species, 16 resident-migrant species, and 5 migrant species. Thirty species of Least Concern and 5 Near Threatened species were recorded in the area during the period. Little egret, Little Cormorant, Purple Moorhen, Purple Heron, Little Grebe, Spot billed duck, Black crowned Night Heron, Indian Pond Heron, Common coot, River tern, White breasted Kingfisher, and Whiskered Tern were the most abundant resident and migrant species foundin the Periyakulam wetlands.
Foraging strategy and prey-handling time in White-bellied heron Ardea insignis in Namdapha Tiger Reserve, Arunachal Pradesh, India
Mondal, Himadri Sekhar; Maheswaran, Gopinathan
Zoological Survey of India, M-Block, New Alipore, Kolkata-700053, West Bengal, India
The White-bellied heron (Ardea insignis) is Critically Endangered and no detailed studies have been carried out prior to ours across the species’ range. Studies on the foraging behavior and prey handling time of the heron were undertaken from November 2013 to March 2016 in Namdapha. Focal animal sampling was used to record the foraging behaviour of herons. The major fish species of the reserve were used for length (cm) and wet weight (g) estimates. The White-bellied heron is a visual forager and adapts to forage amidst fast-flowing freshwater rivers in India, especially within Namdapha. Number of foraging attempts and number of fish caught while ‘facing water current’ (FWC) and ‘against water current’ (AWC) was studied during different months in three seasons. Though the mean number of foraging attempts during FWC (Mean±SD;0.80±1.04)and AWC (0.85±1.05) did not vary significantly (t=-0.63; d.f. 558; n.s.) during three seasons, the number of fish caught did (FWC: 0.34±0.60) (AWC: 0.26±0.51; t=-2.13; d.f, 558; p<0.03). The fish eaten ranged in length from 3-60 cm (mean±SD) (12.9±6.8; n=335) in Namdapha. Fishes of 7 cm were more (11.1%) in the diet of WBH followed by 25 (9%) and 26 cm (9%). As the size of the fish increased in the diet of WBH corresponding prey-handling time also increased.
Status assessment and population trends of the Madagascar Pond Heron, Ardeola idea (Hartlaub, 1860)
Rabarisoa, Rivo1; Ramanampamonjy, Julien R.1; Razafindrajao, Felix2; De Rolland, Lily3
1 Asity Madagascar, BP 1074, Antananarivo 101, Madagascar; 2 Durrell Wildlife Conservation; 3 The Peregrine Fund
The Madagascar Pond Heron, Ardeola idea, is a migratory species breeds exclusively in Madagascar and related islands such as Europa, Aldabra, Mayotte and Comoros. Changes in the population of this species were investigated over the last 20 years through literature reviews, field monitoring and surveys undertaken from 1993 to 2016. Data from 108 localities including the seven known breeding sites were collected and analyzed for the species population assessment. The species occurs to all types of wetlands including lakes, ponds, marshes, rivers, mangroves and also rice field. During the non-breeding season, May-October, birds migrate to eastern and central Africa but some population, with 911 records, remain in Madagascar spent austral winter. Data shows that the current population is evaluated at 2,200 breeding birds remaining into its entire breeding areas. The populations are rapidly declining particularly at its main breeding sites. The main threats are the habitat destruction, collect of eggs and fledgling birds, predation and disturbance at its breeding sites.
Current status of the Madagascar Heron, Ardea humbloti (Milne-Edwards and Grandidier, 1885) in Madagascar
Rabarisoa, Rivo1; Razafindrajao, Felix2; De Rolland, Lily3
1 Asity Madagascar, BP 1074, Antananarivo 101Madagascar; 2 Durrell Wildlife Conservation.; 3 The Peregrine Fund.rivo.
The Madagascar Heron, Ardea humbloti, is endemic to Madagascar and Comoros. The species breeds in Madagascar, with recent records in Mayotte.Distribution and status of the species was investigated over the last 23 years through literature reviews and surveys undertaken from 1993 to 2016 in Madagascar. During this period, 374 records from 108 localities were collected and analyzed. Bird occurs to all type of wetlands habitat with higher concentration recorded to site along the coastal area in western Madagascar; Mangoky Ihotry wetland complex (+100 individuals), Tsiribihina River & delta (+20 individual), Manambolomaty Lakes complex (+300 individual), Baly Bay wetlands (+100 individual), and Mahavavy Kinkony wetland complex (+40 individual). The current population was evaluated at 1,470 individual breeding birds remaining into its entire distribution areas. The population is in declining. The main threats are habitat destruction, disturbance and persecution at its breeding site. Action plan for conservation are needed to preserve this species.
Morphological trade-offs and recursive plumage patterns as indicators of integrated evolutionary dynamics in the Ardeidae
Riegner, Mark F.
Environmental Studies Program, Prescott College, Prescott, AZ 86301, USA
The ultimate form an organism attains is based, in large part, on the rate and timing of developmental trajectories and on compensatory relationships between anatomical structures. For example, there is often an inverse correlation between the size of an organism’s head and the length of its legs. To determine whether a compensatory relationship exists between relative head size and leg length in Ardeidae, I measured skull dimensions (length, width, and height of cranium, and total length including bill) and skeletal limb dimensions (femur, tibiotarsus, and tarsometatarsus) of the 12 North American species as well as 10 other taxa, including the morphologically divergent Cochlearius. In addition, plumage pattern was tabulated across Ardeidae to assess whether an association exists between various patterns and specific morphological traits. My comparisons show that, in general, there is a negative association between proportionate head size and leg length. Ardea species exhibit the smallest relative head size while Cochlearius, Nycticorax, and Nyctanassa have the relatively largest heads. Similarly, Butorides and Ixobrychus have disproportionately large heads compared to leg length, but since their intracranial proportions are average for the family, the relatively large size of the head results from disproportionately short legs. I propose that the long legs of Ardea derive from hypermorphosis while the short legs of Butorides and Ixobrychus are paedomorphic features, which, in compensation, permit the feet to evolve a specialized prehensile function for grasping branches and reeds. Regarding plumage pattern, there are identifiable trends; for example, smaller species and those that forage in enclosed habitats tend toward cryptic patterns. Additionally, specific plumage patterns are not restricted to closely related taxa but recur in diverse genera across Ardeidae.
Prey Consumed by Wading Birds in Mangrove Swamps of Colombia Caribbean Coast
Ruiz-Guerra, Carlos1,2; Echeverry-Galvis, María Á.1
1 Departamento de Ecología y Territorio, Facultad de Estudios Rurales y Ambientales, Transversal 4 #42-00, piso 8 Bogotá, Colombia; 2 Asociación Calidris, Carrera 24 # 4-20, Santiago de Cali, Colombia
Understanding the diet of wading birds can act as an instrument to study relations of key ecosystems, such as mangroves. Although the diet of various species has been studied due to their tendency to regurgitate upon capture, for many species their food requirements are still unknown. In 2015, from May until December, we studied the diet composition of wading birds in the best-conserved area of mangrove swamps of Northwestern Caribbean coast of Colombia, This study encompass both non-breeding season for Little Blue Heron, Tricolored Heron and Snowy Egret and breeding season for Agami Heron, Cocoi Heron, Boat Billed Heron and Bare Throated Tiger Heron. For all species, except Little Blue Heron, fish was the most common prey, with guppies as the most common item followed by gambusia; even with in reduced samples for some egret species. Samples from Agami Heron and Bare-Throated Tiger Heron were too difficult to obtaining for both adults and nestlings, proving them still the most poorly known diet of wading birds in the area and in general in the Neotropic. Based on this information, we are assembled a trophic network to better understand the role of wading birds in mangrove areas, and how this can be affected or not due to human intervention. We found out two families of fish, Engraulidae and Poecilidae are the most vulnerable resource of the trophic network, which are used as bait by artisanal fishermen and are also preys eaten by wading birds.
Herons in Colombia; status, knowledge gaps and conservation
Asociación Calidris, Carrera 24 # 4-20, Santiago de Cali, Colombia
Colombia is host of 24 species of herons including both migrant and breeding populations. This significant diversity is due to the geographical position that includes a large number of biomes and wetlands complex in the country, such as Orinoco floodplains, Amazon forests, and Pacific and Caribbean coasts. However, few studies have focused on herons in this country and information about genera Syrigma, Tigrisoma, Agamia, Cochlearius, Ixobrychus, Botaurus and Zebrilus is scarce. The vast majority of this species inhabits on mangrove forest, swampy forest and natural grasslands that are being profoundly transformed by legal and illegal mining, urbanization, agriculture development, and infrastructure. We made and analysis of risks for herons in Colombia, crossing species distribution and national scale threads. As a result of this analysis we propose two heron species to be included in the Colombian Red List: Zig-zag heron and Agami heron.
A hop, skip and a jump: The use of long-term banding datato understand movement and survivorship of the Reddish Egret in Texas and Mexico
Rylander, Rebekah J.1; Palacios, Eduardo2; Duarte, Adam3; Green, M. Clay4
1 Aquatic Resources PhD Program, Texas State University, San Marcos, Texas, USA; 2 Centro de Investigación, Científica y de Educación Superior de Ensenada, (CICESE), Unidad La Paz, BCS, México; 3 Department of Fisheries and Wildlife, Oregon State University, Corvallis, Oregon, USA; 4 Department of Biology, Texas State University, San Marcos, Texas, USA
Understanding a species’ dispersal and movement behavior is critical when developing a conservation strategy at local and global scales. Without knowledge of the species’ life history in regards to movement patterns, it can be difficult to create an appropriate management plan across and within its range, especially if the species in question travels great distances. The Reddish Egret (Egretta rufescens, REEG) is a medium-sized heron that displays plumage-dimorphism (dark and white). Its range is along the coast of the Gulf of Mexico, Pacific coast of Mexico, as well as portions of Central American and the Caribbean islands. The primary objective of this research was to estimate survivorship of juvenile REEG through long-term color banding, as well as analyze the movement ecology of individually marked birds across Texas and Mexico. During the breeding seasons between 2006-2016, we color banded REEGs in Texas/Tamaulipas (n=628), Yucatan (n=105), Chiapas (256), and Baja California Sur (n=220). Using multi-state models, we analyzed the movement between color morphs (dark and white), sexes (when known), and the four target breeding regions mentioned above. Variation in movement measurements and juvenile dispersal behaviors was seen among individual birds and within different regions. Our results also suggest that juvenile REEG suffer from high mortality rates, which leads us to believe that this life stage is possibly a limiting factor on the growth of the population across its range. Therefore,our research targets the importance of long-term color banding, yielding to considerable insight on survivorship and movement of a species. As more band-resight data is gathered, our ongoing research of REEG will hopefully contribute to the conservation of one of North America’s rarest herons.
Response of Avian Communities to Water Management in the Floodplain Grasslands of the Mekong Delta
Senner, Paul1; Silbernagel, Janet1; Barzen, Jeb2
1 Nelson Institute for Environmental Studies, University of Wisconsin-Madison, WI 53703, USA; 2 International Crane Foundation, Baraboo, WI 53913, USA
Critically important to waterbirds, wetlands in the world’s deltas are disappearing at an alarming rate. In locations such as the Vietnamese Mekong Delta, the loss of natural wetlands is almost complete, necessitating research into the restoration and conservation of the remaining remnants. At Tram Chim National Park (TCNP), a restored floodplain grassland in the Mekong Delta, managers use water control structures to conduct season long draw-downs, mimicking the flood pattern of the Mekong River. Few publications have studied avian communities in Southeast Asian floodplain grasslands or evaluated the success of season long draw-downs in maintaining these communities. At TCNP, changes in water management coincided with two surveys of bird and plant communities, one during a period of dry-season water draw-downs (1986-1995) and another following a period where high water levels were maintained year-round (1996-2005). In the spring of 2016, we repeated these surveys to assess the status of TCNP’s floodplain grassland communities following the resumption of draw-downs in 2006. Using results from the previous surveys as a reference, we assessed species turnover and used taxonomic and functional diversity indices to compare the avian and vegetative communities present during different water management regimes. In 2016, we recorded 33 plant species and 2,241 individuals of 46 bird species. We found that grassland plant communities remained stable across all three surveys. In contrast,avian species diversity was lowest when high water levels were maintained year-round. Grassland specialists (e.g., Megalurus sp. and Anthus sp.) were most abundant in draw-down years, while Ardea sp. and Phalacrocorax sp. were more abundant with high water. As currently implemented, dry-season draw-downs are an effective management technique to maintain floodplain grassland communities at TCNP.
Distribution and Temporal Trends of Western Reef Heron (Egretta gularis) Populationsalong the Arabian Gulf Coast, United Arab Emirates
Shah, Junid N.1; Javed, Salim1; Ahmed, Shakeel1; Khan, Shahid B.1; Al Hammadi, Abdullah1; Soorae, Pritipal Singh1; Al Dhaheri, Shaikha Salem1; Durham, Sarah E.2; Green, M. Clay2
1 Terrestrial & Marine Biodiversity Sector, EAD -Abu Dhabi, Abu Dhabi, UAE; 2 Department of Biology, Texas State University, San Marcos, Texas 78666, USA
The Arabian Gulf of the United Arab Emirates contains highly productive mangrove and seagrass habitats that are essential breeding and non-breeding areas for vast numbers of waterbirds. Unfortunately, both mangroves and seagrasses are declining in this area due to anthropogenic disturbances such as pollution, development, and commercial and recreational activities. Due to their dependence on these coastal habitats, waterbirds such as the western reef heron (Egretta gularis) can act as important bioindicators of ecosystem health. The estimated global population size of E. gularisis roughly 10,000 to 100,000 individuals. While populations are considered stable, many important breeding and overwintering areas of the western reef heron remain unprotected. We performed line transect counts at fifteen sites along the Arabian GulfCoast of the U.A.E. monthly from 2006-2015. Using a mixed effects model, we found that counts varied by both month and year independently, suggesting no interaction between the two fixed factors. Our model analysis also suggested a decline in annual abundance by approximately three birds per year, but lacked statistical significance. We found significant inverse relationships between year and count at the Al Aryam and Abu Al Abyad sites, suggesting annual abundance may be decreasing in these areas. Furtherresearch and monitoring of the U.A.E. western reef heron population is recommended due to habitat loss and potential declines of the species.
From Argos to conservation: reporting 4 years of action for the Agami Heron
Stier, Anna
GEPOG Association, 15 Ave. Pasteur, 97300, Cayene, French Guiana
The Agami Heron is ranked 13th among the world’s conservation priority heron species, and 2nd for the Americas. In French Guiana, which holds the world’s largest Agami Heron colony (representing over 95% of the known population), knowledge was lacking until recently to be able to evaluate the threats on the species and develop an effective conservation plan both for French Guiana and throughout its distribution in South and Central America. To achieve this objective, GEPOG (Group for the Study and Protection of Birds in French Guiana) used the results of two years of Argos tracking of 8 individuals and built up a working group under HeronConservation (Agami Heron Working Group -AHWG), including several experts and natural protected area managers across South America and beyond. To be effective to serve conservational purposes, research results need to be set in perspective with local and regional environmental, economic, social and political conditions, before being declined in concrete and adapted actions.
Abundance and distribution patterns of two heron species in multiple agricultural landscapes of south Asia
Sundar, K. S. Gopi1,2; Kittur, Swati2
1 International Crane Foundation, Program SarusScale, E-11376, Shady Lane Road, Baraboo, WI 53913, USA; 2 Nature Conservation Foundation, Cranes and Wetlands Programme, 3076/5, 4th Cross, Gokulam Park, Mysuru 570002, India
Multi-cropped agricultural landscapes in South Asia have recently been discovered to host surprisingly high bird diversity despite a very long history of cultivation and high human density. Nuanced understanding of how herons interact with the seasonal crops and persisting wetlands is absent. Comparisons across landscapes with different levels of cultivation and human densities are also absent. We systematically collected information on seasonal abundance and distribution of two heron species, the Grey Heron Ardea cinerea and the Purple Heron Ardea purpurea, simultaneously in five agricultural landscapes in lowland Nepal and north-central India during 2014-2016. This project is part of a long-term monitoring framework developed to track the status of large waterbirds in south Asia. We also generated detailed wetland maps for each landscape to determine if abundance and distribution of the two heron species was influenced by wetland extent and position. Both heron speciesshowed: (1) significant spatial and temporal variations in both abundance and distribution in all five landscapes, and (2) considerable complexity in their associations with wetlands varying between landscapes and seasons. Agricultural landscapes showed considerable variations in their utility as heron habitat, with strong evidence of cropping patterns, agricultural intensity, and human population density influencing observed patterns. This is the first assessment carried out simultaneously in multiple landscapes and covering all three seasons for herons in agricultural landscapes. We will discuss the conservation implications of the findings, and future research needs.
Making a case for long-term monitoring of large waterbirds inagricultural landscapes: preliminary findings from south Asia
Sundar, K. S. Gopi1,2; Kittur, Swati2
1 International Crane Foundation, Program SarusScale, E-11376, Shady Lane Road, Baraboo, WI 53913, USA; 2 Nature Conservation Foundation, Cranes and Wetlands Programme, 3076/5, 4th Cross, Gokulam Park, Mysuru 570002, India
Scientific attention to large waterbird ecology outside of the United States and Europe, unfortunately, continues to be low. Understanding of large waterbird needs is therefore biased, with literature frequently suggesting that intensively-cultivated agricultural landscapes have poor utility as large waterbird habitat. Assumptions regarding the impact of areas with high human densities and long agricultural histories on waterbird populations drive global status assessments despite lack of empirical evidence. In south Asia, agriculture is the dominant land use with some areas experiencing cultivation for several centuries. In other areas, government policies are driving increases in cultivation with concomitant alterations in land use, particularly wetland persistence. These landscapes remain largely unexplored for biodiversity, with meagre to no information existing on large waterbirds in these areas. In 2012, as part of Program SarusScape,we instituted a systematic, spatially-explicit, landscape-scale, long-term monitoring program focusing on large waterbirds (cranes, storks, ibis, spoonbills and herons) in seven areas in lowland Nepal and north-central India. These areas were chosen to represent variations in cropping history, agricultural intensity, crop species, human population density, rainfall patterns, and wetland persistence. The effort has yielded: (1) new populations of species of global conservation concern (Sarus Crane, Lesser Adjutant, Asian Woolly-neck); (2) novel information on heronry ecology; (3) evidence for landscape-scale effects of croplands on large waterbird distribution; and (4) a high value of agricultural landscapes as habitat for several large waterbird species. Weposit therefore that assumptions regarding status of large waterbird species using land use alone require to be revisited. Evidence-based understanding of the utility of different habitats and land uses to large waterbirds is critical to obtain realistic large waterbird status assessments.
Collective Colony Formation Algorithm
Toquenaga, Yukihiko1; Tornero, Luis Carrasco2; Mashiko, Miyuki3; Fasola, Fasola, Mauro4
1 University of Tsukuba, Japan; 2 Center for Ecology and Hydrology, UK; 3 Wildlife Management Office, Inc., Japan; 4 University of Pavia, Italy
Colonies of breeding herons and egrets should be located so as to maximize individual resource intake. Food spectra of these species are relatively strict, and their colony sites are restricted to wood and wetlands, so it is relatively easy to specify their habitat requirements. But simple calculation of availability of food and nesting habitats often fails to explain why colonies were formed at specific locations. We tackled this problem by a different analysis of heronry locations, using long-term data for large study areas in Italy and Japan. First we incorporated not only favorable resource sites, such as nesting and foraging sites, but also unfavorable sites, such as urban and bare ground, for evaluating probability of colony formation. Second we incorporated site fidelity, or tendency to reuse the same places as colonial sites. The second aspect was inspired by the fact that nearly two thirds of colonies formed in the past 30 years in our study area in Japan, and in the past 43 years in Italy, were located exactly at the same or at nearby sites. We applied the random forest analysis to create the collective brain, or colony locating algorithms. Their collective brains show that they used specific scale and land use combinations for colony formation, avoiding urban or bare fields at the 10 km scale, but favoring ever green forest at the 4 km scale. Importance of site fidelity constantly increased after 1995 in Italy, and after 2005 in Japan. We discuss the change in colony formation algorithms according to the change of member species in colonies.
Potential Impacts in heron populations due to scenarios of climate change in South America: focus in Paraguay and bordering countries
1 Guyra Paraguay, Parque Ecológico del Rio, BioCentro, Avda. Carlos Bóveda s/n., CC 1132, Viñas Cue, Asunción, Paraguay; 2 Facultad Politécnica, Universidad Nacional de Asunción, San Lorenzo, Paraguay
Climate change affects ecosystems, habitats and species with increasing velocity and continuity. Climate change is expected to cause shifts in the geographic distribution of species worldwide as species track their optimal habitat which will likely shift as a result of rearrangement of climate zones. Water and wetlands are very linked to heron populations and severe extreme conditions such as drought and flooding may affect these populations in light of the different future scenarios related to shifts in temperature and rainfall. We here analyze the historical information on climate for Paraguay and the distributional ranges for the 14 Ardeidae species occurring in the Paraguayan territory and their contribution to the 57 country’s Important Bird and Biodiversity Areas (IBAs). Though most of the Paraguayan heron species may not be severely affected by the different climate change scenarios, this contribution highlight some of the threats and identify strategies for creating conditions towards more resilient heron populations which will serve to decision makers for improving their response andsupport for natural resource management.
——— Poster Presentation ———
Importance of Costa Rican Conservation Areas and IBAS for Ardeidae conservation and management
Alvarado Quesada, Ghisselle M.
National Museum of Costa Rica
Costa Rica has 350 documented wetlands (350,000 ha) in the National Wetland Inventory; all them administrated by Conservation Areas (Environmental Ministry). However the diversity of more of them is poorly known (except Ramsar Sites). Ardeidae species liveand move along the country. I used 55,000 electronic recordsfrom the National Museum and Global biodiversity information facility (GBIF) to create Conservation Areas and IBAS ardeidae species lists and prioritize them by species diversity, vulnerability and future needs. I applied ecological niche modelling for Costa Rica threatened or sensible species: Agami Heron, Least Bittern, Pinnated Bittern and Rufescent Tiger-Heron.
Great Egret roosting dynamics along the North Carolina coast
Andersson, Annika G.; Kornegay, M. Edye; Schweitzer, Sara H.
North Carolina Wildlife Resources Commission, New Bern, NC 28562, USA
To understand more about the habits, habitat preferences, species composition, abundance trends and overall importance of Great Egret (Ardea alba) roosting sites, we monitored eight roosts along the North Carolina Coast from 2015–2016. These roost locations, in Carteret County and at Lake Mattamuskeet National Wildlife Refuge, were obtained through reported observations of tagged egrets from concerned citizens and from a separate radio-telemetry project. Only two of the eight roosting sites have been used consistently by Great Egrets throughout the year; these roosts are on suburban, privately-owned land and are at risk of development or other manipulation. Three roosts are used on a short-term basis (≤3 consecutive months), two sites have been deemed inactive, and we lost access to another. Roost size ranged from 0–259 Great Egrets. High counts of other colonial waterbirds at these roosts include 6 Snowy Egrets (Egretta thula), 46 Cattle Egrets (Bubulcus ibis), 20 Black-crowned Night Herons (Nycticorax nycticorax), 15 Little Blue Herons (Egretta caerulea), 7 Green Herons (Butorides virescens), 7 Great Blue Herons (Ardea herodias), 1 Tri-colored Heron (Egretta tricolor), 383 White Ibis (Eudocimus albus), 18 Glossy Ibis (Plegadis falcinellus) and 1097 Double-crested Cormorants (Phalacrocorax auritus). Our data collection is ongoing and will supplement other roost monitoring projects in North America.
The correlation between prey item distribution and Reddish Egret (Egretta rufescens) foraging behavior
1 Department of Biology, Texas State University, San Marcos, Texas 78666, USA; 2 Department of Geography, Texas State University, San Marcos, Texas 78666, USA
The Reddish Egret is a coastal wading bird with an IUCN conservation status of near threatened, and is listed as threatened in Texas where much of its breeding population occurs. We utilized a Texas Parks and Wildlife Department coastal fisheries database to examine spatial and temporal factors driving prey community structure in the Laguna Madre of Texas. We divided the Laguna Madre into three regional zones: Upper Laguna, Landcut, and Lower Laguna. The fisheries data was collected from 1976-2014 and consisted of multiple seine hauls each month. Environmental data including salinity, turbidity, and temperature were also collected corresponding to each seine haul collection event. We limited the fisheries data to only include those species either known to be prey of Reddish Egrets or were members of the same genus as known prey species. The dataset was also broken into breeding, post-breeding, and migration-wintering seasons based on Reddish Egret life history. We used multivariate analysis to determine what biotic and abiotic factors influenced prey species distribution. This multivariate analyses showed prey varied by both season and region within the Texas Laguna Madre. Future analysis will examine the relationship of prey distribution to known Reddish Egret colony site location and nest initiation date.
Population Viability of the Reddish Egret (Egretta rufescens) in Texas: An Analysis of Management Actions and Implications
Durham, Sarah; Green, M. Clay
Wildlife Ecology Program, Department of Biology, Texas State University, San Marcos, TX 78666, USA
Reddish egrets are a threatened waterbird species that inhabit the Gulf Coast of the U.S. and Mexico, as well as, the Bahamas, Cuba, the Mexican Pacific Coast, and the Yucatan peninsula. The plume trade of the late 1800s drastically reduced global population numbers of reddish egrets. By the 20th century, the species was decimated and possibly extirpated in many parts of its range. While much of the historical range has been recolonized, the reddish egret remains North America’s least abundant heron species. An estimated one-third to one-half of the global reddish egret population occurs in the United States, with Texas having approximately 75% of the breeding pairs. While egret population numbers may be increasing throughout portions of the range, many factors continue to threaten the persistence of the species. Population viability analyses (PVAs) are a common method of predicting a species’ persistence into some future time. The purpose of developing a population viability analysis for E. rufescensis to identify possible factors impeding the growth of Texas populations. By assessing the relative threat of each contributing factor and identifying vulnerable life stages, a robust PVA can estimate how different management actions will affect population demographics. The outcome of this analysis will help guide the management of Texas populations of E. rufescens. Using population and demographic data from the Texas population of E. rufescens, our model will serve as the backbone for the creation of a range-wide PVA.
Status and Conservation of White-eared Night Heron in Vietnam
Hung, Le Manh1; Manh, Vuong Tien2
1 Institute of Ecology and Biological Resources, Vietnam Academy of Science and Technology, 18 Hoang Quoc Viet st, Caugiay dis, Hanoi, Vietnam; 2 Vietnam CITES Management Authority, 02 Ngoc Ha st, Ba Dinh dist, Hanoi, Vietnam
For the last ten years, several works have been done for the little known and endangered White-eared Night Heron Gorsachius magnificus in Northern Vietnam. From direct field surveys and interview the local hunters, we confirmed the records of this species at three different sites in two provinces in Northern Vietnam including (Cho Don and Ba Be districts of Bac Kan province), Trung Khanh district of Cao Bang province. We have also confirmed the occurrence of 8 breeding pairs and estimated the population size in Northern Vietnam is under 100 individuals. The main threats to the species have been identified including habitat loss and hunting. Several conservation works have been done such as setting up the nest protecting group at Xuan Lac and Ba Be sites and the species action plan have been made. Particularly, this species was included in the 160 Degree of Vietnam Government in 2013 which is used as the mainreference for Vietnam penal code and Biodiversity Law.
Changes in populations of colonial herons and egrets in Japan
Mashiko, Miyuki1,2; Toquenaga, Yukihiko1
1 University of Tsukuba. 1-1-1 Tennoudai, Tsukuba, Ibaraki 305-8572, Japan; 2 Current address: Wildlife Management Office, Inc. 1-10-13 Oyamagaoka, Machida, Tokyo 194-0215, Japan
Rice is the most important crop in Japan, and large areas of rice paddies support large populations of colonial herons and egrets. Japanese people have traditionally worshiped herons and egrets as gods protecting crops from pest bugs and locusts. But population sizes of herons and egrets are thought to be declining after World War II due to the decline of areas of rice fields. Here we report colony dynamics around Ibaraki Prefecture, eastern Japan. First, 10-year population dynamics from 2002to 2011 were addressed by censuses based on a combination of aerial photography and ground surveys. Population changes differed among the six constituent species: Great Egret (Ardea alba) and Black-crowned Night Heron (Nycticorax nycticorax) remained relatively constant, while Grey Heron (A. cinerea) and Intermediate Egret (Egretta intermedia) increased, but Little Egret (E. garzetta) and Cattle Egret (Bubulcus ibis) decreased. The marked increase of the Grey Herons contributed to the increasing temporal variation in colony size and in species composition ratio. Second, whether the addition of Grey Herons has affected colony persistence was examined. After the increase of Grey Herons in 2005, colonies with Grey Herons had a greater propensity to persist. Grey Herons began to take the initiative in establishing colonies, and other species began to follow them in colony site selection. The expansion of Grey Herons into mixed-species colonies has promoted the persistence of colonies, and local populations of colonial herons and egrets seemed to gain the benefit of colony sustainability.
Distribution and abundance of egret and night-heron species in Pennsylvania, past and present
Master, Terry L.
Department of Biological Sciences, East Stroudsburg University of Pennsylvania, East Stroudsburg, PA 18301, USA
Of the egret and night-heron species inhabiting Pennsylvania before the turn of the 20thcentury, only the Black-crowned Night-Heron was reported reasonably often as a breeding species. Yellow-crowned Night-Herons and Great Egrets were recognized only as stragglers and post breeding wanderers. At the turn of the 20thcentury, a few nests were reported in southeastern Pennsylvania, also the focus of Black-crowned Night-Heron nesting at the time. Nests of all three species increased in number through the mid 20thcentury, likely a response to recovery from plume hunting and subsequent range expansion. At this time major colony location switched to the lower Susquehanna Valley and environsfrom the southeast, perhaps due to increasing development in this most populated corner of Pennsylvania. In the mid 20thcentury, the three species were joined by relatively short-lived colonies of Snowy and Cattle Egrets, also nesting in the lower Susquehanna Valley. Beginning in the early 1970s, colony locations and numbers were documented routinely and, with the advent of two state-wide atlas efforts spanning 25 years and eBird, a continuous and accurate record of population status has been available since then. Interestingly, throughout this entire period, few individuals and almost no nesting have occurred in the western half of the state. The number of colonies and current population levels of all three species warrant endangered status within the state.
Mitochondrial and Nuclear Phylogenies of the Herons
1 Department of Biology, University of Miami; 2 Musuem of Natural Science, Louisiana State University; 3 University of Texas El Paso; 4 Department of Organismic and Evolutionary Biology, Harvard University
Systematic relationships of the herons (Aves: Ardeidae) are of strong interest to ornithologists. Here we present phylogenies derived from both mitochondrial (cytochrome b) DNA and next-generation sequencing of nuclear DNA, encompassing most of the world's heron species. Phylogenies from both genomic regions were largely concordant with similar structures confirming the monophyly of five main heron groups: (1) Cochlearius (Tigriornis, Tigrisoma); (2) Agamia; (3) Zebrilus (Ixobrychus, Botaurus); (4) Syrigma, Pilherodias, (Egretta), (5) (Butorides, Ardeola) (Ardea, Casmerodius, Mesophoyx, Bubulcus). The night-herons Nycticorax, Nyctanassa, and Gorsachius appear to be polyphyletic.
Effects of hydroelectric generation on foraging in Great Blue Herons (Ardea herodias)
Scarlett, Todd L.
University of South Carolina Lancaster, Lancaster, SC 29720, USA
Tailwater streams below hydroelectric dams are subject to drastic changes in water depth and velocity over short periods of time. The effects of altered flow regimes on macroinvertebrates and fish have received significant study, but the effects on other wildlife have received virtually no attention at all. I examined the effects of altered flow regimes on foraging success and bird abundance of great blue herons (Ardea herodias) at the Lake Wylie Dam near Rockhill, South Carolina, USA. Heron foraging was observed from the dam structure to a distance of 850 m downstream of the dam. During periods of no generation foraging rates were highest at shallow areas 500-850 m below the dam and lowest near the dam. During generation foraging rates were highest immediately downstream of the dam near the outflow of the generators and foraging in the shallows downstream was very low. Censuses conducted every thirty minutes during foraging observations show that significantly more herons were present near the dam during generation than in periods of no generation. Conversely, in the shallow areas 500-850 m below the dam, more heronswere present during periods of no generation than during generation, though the differences at this location were not significant. Great blue herons generally forage solitarily but hydroelectric dams concentrate foraging great blue herons into much greater densities than they are found elsewhere. This may impact breeding colony size and the distance travelled between foraging and nesting sites, particularly during the breeding season when adults are feeding young.
Survival and movements of Black-crowned Night-Herons in Lake Erie, Ohio
Stein, Kristie; Tonra, Christopher
The Ohio State University, Columbus, OH, USA
Located 9 miles due north of the Ohio mainland, West Sister Island is home to 40% of the nesting herons and egrets in the U.S. Great Lakes. One such species of heron, Black-crowned Night-Heron, has experienced a decline in breeding pairs from 3,000 in 1977 to 387 pairs in 1999 and is currently listed as threatened in Ohio (Hothem et al. 2000). Understanding factors influencing movements of individuals across spatial and temporal scales is critical to the preservation of populations threatened by environmental change. Using this information, in accordance with survivorship estimates, is important in understanding population dynamics and the conservation needs of species. To estimate post-fledging survival and examine dispersal patterns, Black-crowned Night-Herons (n=15 fledglings) were marked with coded nanotags in Lake Erie, Ohio, in 2015. Adult night-herons (n=5) were captured at local marinas near public-use fish-cleaning stations using a baited woosh net technique. In combination with active hand tracking, automated telemetry towers were used to passively monitor movement activity of marked birds. During the 2016 nesting season, we will use the same techniques to deploy coded nanotags (n=60) and ARGOS satellite transmitters (n=15 adults). I will present preliminary results on fledging rates, survival, recruitment, and movements of juvenile and adult Black-crowned Night-Herons. Incorporating knowledge of night-heron movement and population demographics may help guide future management decisions to maintain a viable population.
ARGOS tracking to understand the ecology and behavior of Agami Herons
Stier, Anna
GEPOG Association, 15 Ave. Pasteur, 97300 Cayene, French Guiana
The Agami Heron is ranked 13th among the world’s conservation priority heron species, and 2nd for the Americas. Until recently, nothing was known about the feeding grounds of this species during the breeding season, or about areas used during the nonbreeding season. In French Guiana, which holds the world’s largest Agami Heron colony (representing over 95% of the known population), the location and characterization of these habitats as well as the identification of the geographic location and routes travelled by breeding and non-breeding individuals has been crucial to evaluate the threats on the species and develop an effective conservation action plan both in French Guiana and throughout its distribution in South and Central America. To achieve this objective, GEPOG (Group for the Study and Protection of Birds in French Guiana) tracked 8 Agami Herons in 2012 and 2013 via the European LIFE+ Cap DOM program. Data from 4 individuals show that this species migrates north and south along the coast (Brazil, Suriname and Venezuela) and is able to cover up to 1,300 km within two months, including several stop-overs. Sizes and habitats of the breeding season home ranges are determined.
Seasonal variation of Reddish Egret movement along the Gulf of Mexico
Walker, Isabelle; Green, M. Clay
Department of Biology, Texas State University, 601 University Drive, San Marcos, TX 78666, USA
Understanding seasonal variation in waterbird movement is essential for conservation management of key habitats used during breeding, post-breeding and wintering seasons. Reddish egrets (Egretta rufescens) are a near threatened waterbird species with the breeding population primarily occurring along the Texas coast. There has been limited knowledge on their movement especially changes in movement from juvenile dispersal to breeding adult. In this study, we estimate seasonal variation in home range size and site fidelity of reddish egrets. Data was analyzed from five years of satellite telemetry data collected during 2010-2015 from 25 individuals along the Texas Coast in the Laguna Madre. Our results show individual home range sizes were the largest during the 2010 breeding season 3,825 km2 (n=20) and decreased to 145 km2 (n=19) in the 2010 winter season. Home range size varied between age and season with an overall decrease during the study. During 2013-2015, home range size significantly decreased to <3 km2 suggesting further evidence for weak long distance migration in this species. Of the remaining tagged birds from 2013-2015, there was no evidence of site fidelity to the natal colony. All of these birds settled in the southern tip of Texas and the Laguna Madre y Delta del Río with the largest dispersal of 165 km. Our study is the first study to track individual Reddish Egret movement from juvenile age class to breeding age class.
Habitat characteristics and seasonal numbers of Great Egrets at 70 roost sites in the area of Southern Ontario, Canada
Weseloh, D. V. Chip1; Moore, Dave2; Knezevic, Tina1
1 Canadian Wildlife Service, 4905 Dufferin St., Toronto, Ontario M3H 5T4, Canada; 2 Canada Centre for Inland Waters, Box 5050, Burlington, Ontario L7R 4A6, Canada
Roosting sites of Great Egrets (Ardea alba) away from breeding colonies are little known and studied. Our objective was to identify roosting sites,their size and habitat, annual usage and rarity, and to answer the question of whether roost sites should be Important Bird Areas (IBAs). We searched for and monitored roosting sites annually, 2008-15; 70 confirmed roosting sites were located in southern Ontario and adjacent U.S. states; 18 other sites were suspected of being roosts. All roosts were located in wetlands where microhabitat included: adjacent to water in trees on mainland (40%) or islands (11%), shallow water or mudflats (25%), trees in water (11%), bushes in water (7%) and herbaceous vegetation in water (6%). The most common size categories were 6-15 birds (30%) and 16-40 birds (24%) but individual roosts ranged in size from 1 to 816 birds. Most roosts consisted of a single site in a wetland,however, at 38% of the wetlands, egrets roosted sequentially at 2 or more sites within the wetland, i.e. they moved to a 2nd site within the wetland, often within the same season. They often moved from trees to mudflats as water levels receded. Roosts usually became active during mid-June to early August, had peak numbers in late August-early September and declined in numbers during late September–late October. Not all roosts are worthy of IBA designation.
Long Term monitoring of egrets and herons in Hong Kong (1989–present)
Wong, L. C. Captain; Woo, C. K. Luke; Wong, Y. P. Josephine; Fung, C. L. Louis; Tam, Y. S.; Young, Llewellyn
The Hong Kong Bird Watching Society. 7C, V Ga Building, 532 Castle Peak Road, LaiChi Kwok, Kowloon, Hong Kong
Systematic, territorial-wide monitoring of egrets and herons has been conducted in Hong Kong since 1989. Through the population change, this monitoring could reveal how egrets and herons make use of fragmented wetlands and adapt dynamic change in urbanized landscapes. In recent years, the annual population of five species (Great (Ardea alba), Little (Egretta garzetta) and Cattle Egrets (Bubulcus coromandus), Black-crowned Night Heron (Nycticorax nycticorax) and Chinese Pond Heron (Ardeola bacchus)) fluctuated around 1,000-1,400 nests, with Little Egret and Chinese Pond Heron were the dominant species. The Great Egret exhibited a long term increasing trend (from zero in 1989 to 283 nests in 2015, while Cattle Egret showed a long term decline in the number of nests (from 118 nests in 1989 to 54 nests in 2015). Although pond type wetlands, and farmlands in Hong Kong were degraded in terms of fragmentation and shrinking in area during the period, both the total number of nests and the number of colonies in 2015 are higher than 1989. This may imply that nesting birds tend to split into more small colonies, and these coloniescould be situated in close proximity to feeding habitats. This may help reducing energetic costs. Also, their feeding habitat use pattern may shift to coastal wetlands in order compensate to the loss of pond type wetlands and farmlands. Pollution control in coastal areas, and better management of protected wetlands (for instance the Mai Po Nature Reserve and Hong Kong Wetland Park) may also provide prime feeding habitats although the wetland area was shrinking.
HeronryMAP:Africa - Mapping the distribution and status of breeding sites of Ardeids and other colonial waterbirds in Africa #
Douglas M. HarebottleDepartment of Biological and Agricultural Sciences, School of Natural and Applied Sciences, Sol Plaatje University,
Private Bag X5008, Kimberley, South Africa;
Abstract
Heronries in Africa are poorly studied and many data gaps are evident in terms of occurrence, species composition and productivity of these colonial breeding sites. This paper introduces HeronryMAP:Africa, a citizen-science project started in 2014 that aims to systematically collect long-term data on location, size and composition, site fidelity, longevity and conflict scenarios of heronries in Africa. Preliminary results are presented for current and historical sites sourced over a three year period (2014-2016). Three hundred and thirty-six colony sites were identified and mapped in 14 (25.9%) African countries; 72.6% of sites have no formal protection, 18.8% were subject to at least one human conflict scenario with ‘cutting of trees’ and ‘removal of trees’ being the most common human disturbances. A first, but presumably grossly underestimated total of 35,000 breeding pairs of colonial waterbirds in Africa is provided from available data. No species-specific nest data are given due to the tendency to report total nest numbers in mixed colonies rather than species-specific numbers. The study revealed a general paucity of data for heronries in Africa (there was no response from 74.1% of African countries), but also the challenges faced in collecting adequate scientific data for these sites. It did, however, show how citizen-science can make significant contributions to research projects that are poorly funded or have limited resources. Human-wildlife conflicts were highlighted as an area that is poorly understood for heronries but has important conservation outcomes. Future objectives include identification of species composition, assessment of priority sites, identification of conservation action for colonies under threat and production of an Atlas of African Heronries.
# This paper was presented at the 1st Herons of the World Symposium at the 40th Anniversary Meeting of the Waterbird Society at New Bern, North Carolina, USA, 21-23 September 2016. Other papers from that Symposium have appeared in past (or will appear in future) issues of the Journal of Heron Biology and Conservation, and Waterbirds.
Introduction
Waterbirds that breed communally in freshwater or coastal systems are spread across eight bird families: Phalacrocoracidae, Anhingidae, Pelecanidae, Ardeidae, Ciconiidae, Threskiornithidae, Phoenicopteridae and Laridae (Perennou et al. 1996, Clements et al. 2017). Most of these waterbirds breed in large colonies, either loosely or in close association; however, some species are solitary nesters, e.g. Goliath Heron (Ardea goliath) and White-backed Night Heron (Gorsachius leuconotus) (Hancock and Kushlan 1984, del Hoyo et al. 1992). Colonies may be largely discrete (e.g., pelicans, gulls, terns and cormorants) or mixed (e.g., ibises, herons, egrets and spoonbills) (Hancock and Kushlan 1984, Perennou et al. 1996). However, some species such as the White-breasted Cormorant (Phalacrocorax lucida) and Reed Cormorant (Microcarbo africanus) are also known to frequently nest in extensively mixed flocks with Ardeids and Threskiornithids (DMH pers. obs.). The term ‘heronry’ usually refers to breeding sites where Ardeid species nest in mixed colonies (British Trust for Ornithology 2018); however, for the purpose of this paper, I will use this term to refer to breeding colonies for the colonial species concerned.
Due to their conspicuous behavior, abundance and often socio-economic and ecological impacts, the general distribution and basic biology of most of these taxa have been well studied globally (Hancock and Kushlan 1984, Brooke and Birkhead 1991, Kushlan and Hafner 2000, Kushlan and Hancock 2005). Continentally, studies are generally well distributed: in Europe (Hafner and Fasola 1997, Marchant et al. 2004, British Trust for Ornithology 2018), Asia (Hong Kong Bird Watching Society 2016, Mashiko and Toquenaga 2018, Matsunaga 2018) North America (Gawlik et al. 1998, Spies and Weingartner 2007, Maccarone et al. 2010, Rush et al. 2015, Cox et al. 2017), South America (Kushlan et al. 2002, Stier 2018, Yanosky 2018) and Australia (Maddock and Baxter 1991, Richardson et al. 2001, McKilligan 2005). However, gaps do exist and in Africa, information on the status and distribution of heronries is severely lacking; it is limited mainly to Southern and Eastern Africa (Tarboton 1977, Underhill et al. 2009, Turner 2011, Kopij 2014). Some data have been collected through atlas projects (Tanzania Bird Atlas; N. Baker in litt.), waterbird surveys (Botswana; Tyler 2001, Madagascar; Wetlands International 2012, Dodman 2014, Rabarisoa et al. (in review)) and some dedicated efforts of individual researchers (Turner 2002, J. Agutu unpubl. data, C. Barlow in litt.). However, most of these studies were short-term or of an irregular nature. Currently, only a single long-term monitoring program (1993 to present) for heronries in Africa is known to the author (Rabarisoa et al. (in review)). As a result, there is a gap in the knowledge of the importance of these African sites in terms of location, species composition, abundance, breeding productivity and site management (Perennou et al.1996, Kushlan et al. 2002).
HeronryMAP:Africa was born out of a heron banding project that started in 2002 in Cape Town, South Africa (Harebottle and Gibbs 2004, 2006) and the general paucity and limited nature of information on heronries in South Africa (Tarboton 1977, Perennou et al. 1996, Veen et al. 2011). The project was officially launched on 1 August 2014 through social media with the creation of a web page via Facebook - ‘HeronryMAP:Africa’.
The objective of this paper is to introduce HeronryMAP:Africa as a continent-wide, citizen-science based monitoring project for African heronries; preliminary results on the status and distribution of current and historical heronries in Africa are presented for 31 species (Appendix 1) and gaps in research and conservation interventions are identified and discussed.
Appendix 1. The 31 colonial waterbird species that were considered in determining colonial sites for this study. Nomenclature after Gill and Donsker (2018); global status taken from IUCN (2017). LC = Least Concern, VU = Vulnerable, EN = Endangered. Country codes: BW – Botswana, CV – Cape Verde, GM – The Gambia, KE – Kenya, LS – Lesotho, MG – Madagascar, ML – Mali, NA – Namibia, SZ – Eswatini (formerly Swaziland), TZ – Tanzania, UG – Uganda, ZA – South Africa, ZW – Zimbabwe. Heronries in Senegal and Mauritania did not report any species specific information and are therefore not included here.
Methods
The study area for this paper was the entire continent of Africa, the island of Madagascar and smaller offshore islands including Cape Verde, Madeira Islands, Zanzibar, Comoros, Sao Tome and Principe. Data were collected from various sources including surveys from ornithologists, heron researchers and bird club members; additional information was sourced from nest record cards (Animal Demography Unit, University of Cape Town; unpubl. data), academic or popular literature and from personal observations. The HeronryMAP:Africa web page was used extensively to request data and collate records and information about heronries throughout Africa, especially South Africa; all researchers and observers were encouraged to post records and upload images of active heronries. Standardized datasheets, available on the site, were provided for participants to use in uploading their data in a standardized format (Appendix 2). Any incidental information relating to breeding sites that was posted directly on the page was transferred to a database.
Appendix 2. Sample data Excel forms that were made available to observers for the collection of heronry data for this study: (a) The first version of the data form that was used in the early part of the study; (b) A streamlined modified version of (a) and is the current version that is used.
Data were grouped into current sites, 2012-2016, which had census data and historical sites, pre-2012 which included sites with census data and those which were reported as active but lacking census data. The reason for selecting 2012 as a cut-off to separate ‘current’ from ‘historical’ heronries is based on heronry dynamics; natural heronry sites generally persist for a few (2-3) years (due to natural variable landscape fluctuations or changes) before being abandoned (Perennou 1996, Underhill et al. 2009), therefore five years (prior to the final year of data gathering for this study, 2016) would be a reasonable amount of time to isolate recent, active colonies from older colonies that may have abandoned preferred sites and moved to other optimal sites.
Additional site protection status information and/or human-related conflict issues were sourced by the author where these were not or could not be provided by the respondent or observer.
Results
Spatial distribution and numbers
A total of 336 heronry sites was mapped from 14 countries across Africa (Table 1, Fig. 1); no distinction was made between colonies being mixed or discrete. Most records (n=238, 70.8%) were from southern Africa. Almost a quarter of the sites (n=73, 21.7%) were located in East Africa (including Madagascar), while 24 sites (7.1%) occurred in West Africa, including the Cape Verde Islands. Only one site (0.3%) was reported for Northern Africa (Mauritania, Table 1). No data were received from central Africa. Nearly half (45.8%) of localities (154/336) were recorded from South Africa; the next largest representations were from Kenya (32 sites), Uganda (29), Botswana (26), Lesotho (22), Zimbabwe (21) and The Gambia (21) (Table 1, Appendix 3).
Table 1. Summary of the status of heronry sites in Africa based on submitted and additionally sourced data to HeronryMAP:Africa. The estimated number of breeding pairs is based on available nest count data for active sites only and should be regarded as preliminary estimates. Regions are based on those defined by the African Union (http://www.west-africa-brief.org/content/en/six-regions-african-union): N = Northern, S = Southern, E = Eastern and W = Western. Data sourced over a three-year period, 2014-2016.
Figure 1. Spatial distribution and protective status of 336 heronry sites throughout Africa based on HeronryMAP:Africa data. Protected sites are those located in formally protected areas; partially protected sites are those which are or form part of Important Bird Areas or Ramsar Sites; Unprotected sites are those sites which are known to occur outside of formally protected areas; Unknown refers to sites for which no information was available to determine protective status. Dashed lines indicate boundaries separating the five regions in Africa (see Table 1). The numerical values refer to the number of sites in each region.
Appendix 3. Photos of selected colonies included in the HeronryMAP:Africa database.
Based on available data from submitted and sourced records for active heronries, a preliminary estimate of 35,000 breeding pairs was calculated from the 319 sites (out of 336 total sites) for which there were numerical data (Table 1); the estimate assumes that all historical sites (i.e. prior to 2012; n=162) have remained active with similar colony sizes that were initially reported. Seventeen sites were shown to be active prior to 2012 but lacked actual nest data. Most survey responders did not indicate any species specific numbers within a heronry; unfortunately, no verified species-specific breeding numbers are yet available in this study.
Protection status
Of the breeding colonies, 72.6% (244/336) were located in unprotected areas; 16.4% (55/336) of the sites were located in formal conservation areas (e.g. national parks or nature reserves) or included in Important Bird Areas and/or Ramsar Sites (Table 1, Fig. 1). The protection status for 37 colonies (11.1%) was ‘Unknown’.
Human-conflict coverage
Of the 336 sites, 81.3% (n=273) did not report any known human-wildlife conflicts; the remaining 63 sites (18.7%) had at least one known conflict (Table 1). From these 63 sites, the most frequently recorded conflicts included ‘cutting of trees’ and ‘removal of trees’. The distribution of human-conflict issues across all sites is given in Fig. 2; the current data indicate that Kenya and South Africa recorded most of the conflict scenarios identified.
Figure 2. Spatial distribution and type of human-wildlife impacts of 336 heronry sites throughout Africa based on HeronryMAP:Africa data. Hunting refers to killing of adult birds at nest sites; Mixed impacts refers to any combinations of known impacts and Unknown refers to colonies where there are no data available on conflict scenarios. Dashed lines indicate boundaries separating the five regions in Africa (see Table 1). The numerical values refer to the number of sites in each region.
Discussion
The results presented here are based on data sourced over a three year period and represent at least an initial attempt to document and quantify the numbers and distribution of heronries in Africa. However, it can be assumed that this is a gross under-estimation and under-representation of the real situation given the number of African countries (40 out of 54) for which no data were submitted in HeronryMAP:Africa but which probably have breeding colonies (see Clancey 1997 for Mozambique; Borrow and Demey 2010 for Ghana; and Redman et al. 2011 for Ethiopia, Eritrea, Djibouti and Somalia). This study also highlighted the challenges in collecting large-scale data on nesting colonial waterbirds across the African continent. Data are often insufficient or difficult to source, particularly when sites are known from personal experience (but not documented) or personal communication but where exact details cannot be ascertained in a timely manner. These gaps, as well as the identification of the species composition and updated status of historical sites with no data available, need to be filled in order to present a clearer and more holistic picture of the status, content and distribution of heronries across Africa. Dodman (2014) does include some information on breeding colonies for some species listed in this study but often locality details are lacking or information is vague. These data will need to be sourced so that these sites can be included in HeronryMAP:Africa and any future reviews of the dataset. Ongoing assessments and conservation measures remain limited without this information. The Heron Specialist Group of the International Union for the Conservation of Nature (IUCN) Species Survival Commission recognizes these gaps and has initiated an effort to establish a list of heron researchers in Africa in order to start building a database of heron researchers and to stimulate further development of research and projects on herons (C. King in litt.).
Although the distribution of documented heronries covers only a small number of African countries (n=14, 25.9%), the majority of records stem from southern Africa, and South Africa in particular. This has largely been due to the strong citizen-science networks in the region and the response of these volunteers to requests for information on heronries in the region. Most of these volunteers used social media to supply relevant information. Eastern Africa is the region which has the second largest number of documented heronries. There have been ongoing efforts there to document and update the status of herons in the region. Turner (2011) provided detailed accounts of the status of 19 Ardeidae in eastern Africa. The Tanzanian Bird Atlas (http://tanzaniabirdatlas.net/start.htm), which has now been underway since 1985, is providing valuable high-quality data for ardeid distributions and seasonality and has recently incorporated mapping active heronries into its volunteer operations (N. Baker in litt.). There are an estimated 50+ sites that have already been documented as part of the Tanzanian Bird Atlas with increasing numbers projected in the next five years (N. Baker in litt.). Western Africa has some data on heronries available through their coordinated waterbird monitoring programs (e.g. waterbird counts in Senegal and Mauritania, Veen et al. 2007) and through ex-patriots stationed in certain countries such as The Gambia (C. Barlow in litt.). Data are severely lacking from the rest of Africa, notably northern and central Africa, where up to nine and 26 species of ardeids, respectively, are known to breed (Hancock and Kushlan 1984, Brown et al. 2002).
This analysis has highlighted that only a small percentage (16.4%) of colonies are located in protected areas where sites can be protected from general human disturbance. Sites located on private land or public open spaces (e.g. parks and gardens) are subject to unpredictable threats such as human disturbance, including destruction of nests, egg predation and cutting or removal of trees. Considering most sites lack formal protection, conservation efforts for colonial waterbirds may need to be focused on private landowners in order to secure breeding sites across Africa and which may include landowner stewardship (https://www.capenature.co.za/care-for-nature/stewardship/) and custodianship programs (Little and Theron 2014).
The results of this study explicitly show that human-wildlife conflict scenarios are generally poorly understood or recognized for colonial waterbirds in the African landscape. Expansion of species and breeding sites into urban, suburban and rural areas often bring them into close contact with people and their associated activities (Telfair et al. 2000). These species/colony-human associations regularly lead to confrontation resulting mainly from the birds’ nesting and breeding activities and guano deposits posing nuisance factors and potential health risks. Many colonies are at risk; some are labelled nuisance sites due to excessive noise of breeding birds and potent guano smell (Grant and Watson 1995, Whittington-Jones 2014), while others are located close to airports or airfields creating potential collisions with aircraft as birds traverse the airfield to and from the colony (A. Froneman in litt.). Consequences of this are that colonies are usually destroyed (either through nest removal or tree cutting) without proper intervention or guidance from relevant authorities. There are no formal regulations or systematic guidelines in place to ensure that these situations are handled in a proper manner. In South Africa, however, Harebottle et al. (2019) have developed national guidelines to assist affected parties and provincial authorities in identifying and mitigating problematic colonies. Similar initiatives in other African countries, particularly where large heronries are under threat from human disturbance, should be considered. Nesting habitat enhancement has been carried out in South Africa (Harrison et al. 2001, Harrison 2005), by building artificial platforms for colonially nesting waterbirds. These have been constructed to replace natural sites that were not being used or were destroyed, and to attract species to breed in new areas. The platforms have been used to varying degrees of success but generally birds respond positively to these artificial nesting structures (Harrison et al. 2001, Harrison 2005). Management and maintenance of the platforms or structures are required to ensure sustainability of breeding populations on an annual basis. Mitigating human-wildlife conflict situations for colonial waterbirds may involve increased focus on constructing artificial breeding sites, particularly where threatened species are present and/or large, natural sites are under increasing threat (Perennou et al. 1996, Harrison et al. 2010).
The breeding pair estimates across all species within each country gleaned from this study should be interpreted cautiously. They are based solely on submitted information, and in light of missing data from other colonies, are gross under-estimations. At best, the figures given in this study should be regarded as an initial attempt to gauge the relative importance of breeding sites and abundance in each country. For colonial waterbirds, numbers of nests per active colony usually relate to breeding success (Perennou et al. 1996). This is driven largely by the number of pairs (within species and across species) that can build nests (nest site availability) and raise chicks. HeronryMAP:Africa will attempt to monitor breeding numbers and output as part of its long-term objectives.
Continued data collection, analysis and site assessments are critical to identify and document additional sites, determine their status and potential productivity, and assess the degree of risk to the future of the sites from habitat loss, climate change, human-wildlife-conflict and other conservation threats. The use of modern technology is crucial to collect high-quality data rapidly; this is important given the real possibility that, in the absence of any formal regulations, large and potentially important colony sites may be subject to disturbance and destruction. In South Africa, the development of mobile applications, such as BirdLasser (https://www.birdlasser.com) allows for project specific data to be collected; BirdLasser incorporates a HeronryMAP:Africa function which prompts users to add additional information (including colony name, breeding status of each species, etc.) when they log any of the species listed in Appendix 1.
Future long-term priorities for HeronryMAP:Africa will be to identify, census and prioritize sites at national and regional levels. This should be based on a set of criteria that will include the number of each species at the colony, conservation status of the site, species of greatest conservation need and threats to the colony. This will focus attention on important colonies and particularly those threatened by human disturbance. In addition, prioritization of heronries will enable conservation authorities to include these nesting sites in national or regional conservation planning programs. Kushlan et al. (2002) and Kushlan (2007) emphasize that where nesting habitat for colonial waterbirds is limiting, these habitats need to be protected and managed effectively to ensure survival of healthy populations. A broader landscape-habitat approach may need to be considered as part of the HeronryMAP:Africa project to ensure habitats and sites can be identified and managed, which may include the need to set up alternative, man-made sites; Perennou et al. (1996) and Kushlan et al. (2002) stress that the importance of artificial nesting sites should not be underestimated. In addition, the impact of climate change on wetland hydrology needs to be investigated as it may affect habitat quality, availability of nest sites and the timing of nesting and migration (Kushlan et al. 2002). Climate change may ultimately impact negatively on priority sites and birds may be forced to source alternative, potentially inferior sites as future breeding colonies. Kushlan (1993) identified colonial waterbirds as effective bioindicators of environmental change and HeronryMAP:Africa could be an effective data source to further elucidate how colonial waterbirds will respond to environmental change.
This study has highlighted the power of citizen-science and the use of social media in creating awareness and garnering biodiversity data for conservation outcomes. McKinley et al. (2017) and Sullivan et al. (2017) both highlight the importance of citizen-science and open access data in providing adequate information for species conservation action and natural resource management. Newman et al. (2017) argue that growth in technologies, particularly mobile applications, has the potential to expand the frontiers of social media and citizen-science to advance scientific research programs; they further elaborate that socio-cultural issues will likely influence citizen-science programs in the future as more biodiversity issues become linked to landscape and habitat changes. This is particularly relevant to this project as breeding sites may increasingly occur in local villages or areas of higher population densities resulting in potential human-wildlife conflict but also in opportunities for local citizen-science participation. In addition, access to social media platforms and technologies may be limited in parts of Africa (e.g. central and northern Africa) and which resulted in a lack of response and hence no data submissions for this study. Ultimately, long-term funding is pivotal to ensure that the HeronryMAP:Africa project can be effectively coordinated and managed into the future, where new or existing technologies for data collection can be implemented and coverage widened to include gap areas.
A long term outcome for HeronryMAP:Africa will be the production of an ‘Atlas of African Heronries’ which can serve as a valuable conservation resource. HeronryMAP:Africa, however, should be seen as an ongoing, long-term initiative to monitoring breeding populations of colonial waterbirds and species as well as implementing conservation action at both local and regional scales. The project also has the potential to provide improved species population estimates at country or regional levels which can contribute to Wetlands International’s Waterbird Population Estimates (Wetlands International 2012).
Acknowledgements
I am grateful to Dalton Gibbs for the inspiration during our early heronry ringing expeditions in Cape Town; this is where thoughts and ideas germinated. To all the people who responded to my requests for information, I am indebted to their input and timely response; without their submissions, data would be severely limited and outputs hampered. A special thanks to Don Turner for allowing access to his data and reports for eastern Africa, and Clive R. Barlow and The Gambian Department of Wildlife for data and information from The Gambia. I am extremely thankful to all the citizen-scientists who have joined the Facebook group, submitted photos, data and information and share a passion for heronries and heronry conservation; without their support this paper would not have been possible. Tabaro Kabanda kindly assisted with the production of the final maps. Lastly, I am indebted to Clay Green, Katsutoshi Matsunaga, Chip Weseloh and two anonymous reviewers for their input and comments that vastly improved the quality of the manuscript.
Literature Cited
Borrow, N. and R. Demey. 2010. The birds of Ghana. Bloomsbury, London, U.K.
British Trust for Ornithology. 2018. Heronries census. [online] Accessed 20 August 2018.
Brooke, M. and T. Birkhead 1991. The Cambridge encyclopedia of ornithology. Cambridge University Press, Cambridge, U.K.
Brown, L. H., E. K Urban and K. B. Newman (eds.). 2002. The birds of Africa. Volume 1. Christopher Helm, London, U.K.
Clancey, P. A. 1997. The birds of southern Mozambique. African Bird Book Publishing, Westville, South Africa.
Clements, J. F., T. S. Schulenberg, M. J. Iliff, D. Roberson, T. A. Fredericks, B. L. Sullivan and C. L. Wood. 2017. The eBird/Clements checklist of birds of the world: v2016. [online] Accessed 25 August 2018.
Cox, W. A., A. Schwarzer, R. Kiltie, A. Paul, M. Racha, K. D. Meyer, J. J. Lorenz, P. E. Frezza, H. Rafferty, G. M. Kent and S. Roebling. 2017. Development of a survey protocol for monitoring Reddish Egrets (Egretta rufescens) in Florida, USA. Waterbirds 40: 334-343.
del Hoyo, J., A. Elliot and J. Sargatal (eds.). 1992. Handbook of birds of the world. Volume 1: Ostrich to Ducks. Lynx Edicions, Barcelona, Spain.
Dodman, T. 2014. Status, estimates and trends of waterbird populations in Africa: AEWA-listed African populations. (CSR6 African populations). Wetlands International Wageningen, The Netherlands.
Gawlik, D. E., R. D. Slack, J. A. Thomas and D. N. Harpole. 1998. Long-term trends in population and community measures of colonial-nesting waterbirds in Galveston Bay Estuary. Colonial Waterbirds 21: 143-151.
Gill, F. and D. Donsker (eds.). 2018. IOC World bird list (v8.1). [online] Accessed 15 November 2018.
Grant, K. R. and J. Watson. 1995. Controlling nuisance egret and heron rookeries in Oklahoma. Great Plains Wildlife Damage Control Workshop Proceedings. Paper 435. [online] Accessed 12 December 2018.
Hafner, H. and M. Fasola. 1997. Long term monitoring and conservation of herons in France and Italy. Colonial Waterbirds 20: 298-305.
Hancock, J. and J. A. Kushlan. 1984. The herons handbook. Croom Helm, London, U.K.
Harebottle, D. M. and D. Gibbs. 2004. Colour-ringed waterbirds in the southwestern Cape. Promerops 258: 9-10.
Harebottle, D. M. and D. Gibbs. 2006. At what age do African Sacred Ibis breed? Promerops 265: 13.
Harebottle, D. M., H. Smit-Robinson and A. Froneman. 2019. National guidelines for interventions that relate to colonial breeding waterbirds causing human-wildlife conflict in South Africa. [online] Accessed 20 June 2019.
Harrison, J. A 2005. Sticks ’n stones: building for birds. Africa - Birds & Birding 10: 52-59.
Harrison, J. A., M. McIver and M. Weyers. 2001. Weyers Islands: a successful design for constructed heronries. Bird Numbers 12: 40-43.
Harrison, J. A., A. J. Williams and M. MacIver. 2010. Breeding site selection by colonial waterbirds given various combinations of constructed or natural alternatives over a 10-year period. Ostrich 81: 197-203.
Hong Kong Bird Watching Society. 2016. Egretry Counts in Hong Kong with particular reference to the Mai Po Inner Deep Bay Ramsar Site – Summer 2016 Report. [online] Accessed 22 March 2018.
IUCN (International Union for the Conservation of Nature). 2017. The IUCN Red list of threatened species. Version 2017-2. [online] Accessed 1 December 2017.
Kopij, G. 2014. Distribution and numbers of breeding cormorants (Phalacrocoracidae) and herons (Ardeidae) in Lesotho. South African Journal of Wildlife Research 44: 189-192.
Kushlan, J. A. 1993. Colonial waterbirds as bioindicators of environmental change. Colonial Waterbirds 16: 223-251.
Kushlan, J. A. 2007. Conserving herons. A conservation action plan for the herons of the world. Heron Specialist Group and Tour de Valat, Arles, France.
Kushlan, J. A. and H. Hafner. 2000. Heron conservation. Academic Press. London, U.K.
Kushlan, J. A. and J. A. Hancock. 2005. The herons. Oxford University Press. New York, New York, U.S.A.
Kushlan, J. A., M. J. Steinkamp, K. C. Parsons, J. Capp, M. A. Cruz, M. Coulter, I. Davidson, L. Dickson, N. Edelson, R. Elliot, R. M. Erwin, S. Hatch, S. Kress, R. Milko, S. Miller, K. Mills, R. Paul, R. Phillips, J. E. Saliva, B. Sydeman, J. Trapp, J. Wheeler and K. Wohl. 2002. Waterbird conservation for the Americas: The North American waterbird conservation plan, version 1. Waterbird Conservation for the Americas, Washington D.C., U.S.A.
Little, I. T. and C. Theron. 2014. Guidelines for custodianship in South Africa, rewarding conservation friendly farming. Unpublished report of the Endangered Wildlife Trust, Johannesburg, South Africa. [online] Accessed 27 June 2019.
Maddock, M. and G. S. Baxter. 1991. Breeding success of egrets related to rainfall: a six-year Australian study. Colonial Waterbirds 20: 133-139.
Maccarone, A. D., J. N. Brzorad and H. M. Stone. 2010. Nest-activity patterns and food-provisioning rates by Great Egrets (Ardea alba). Waterbirds 33: 504-510.
Marchant, J. H., S. N. Freeman, H. Q. P. Crick and L. P. Beaven. 2004. The BTO heronries census of England and Wales 1928-2000: new indices and a comparison of analytical methods. Ibis 146: 323-334.
Mashiko, M. and Y. Toquenaga. 2018. Site fidelity in lineages of mixed-species heron colonies. Waterbirds 41: 355-364.
Matsunaga, K. 2018. Changes of the nesting sites of Grey Herons (Ardea cinerea) in Hokkaido, northern Japan. Journal of Heron Biology and Conservation 3(1): 1-7. [online] Accessed 15 November 2018.
McKilligan N. 2005. Herons, egrets and bitterns. Their biology and conservation in Australia. CSIRO Publishing, Collingwood, Victoria, Australia.
McKinley, D. C., A. J. Miller-Rushing, H. L. Ballard, R. Bonney, H. Brown, S. C. Cook-Patton, D. M. Evans, R. A. French, J. K. Parrish, T. B. Phillips, S. F. Ryan, L. A. Shanley, J. L. Shirk, K. F. Stepenuck, J. F. Weltzin, A. Wiggins, O. D. Boyle, R. D. Briggs, S. F. Chapin, D. A. Hewitt, P. W. Preuss and M. A. Soukup. 2017. Citizen science can improve conservation science, natural resource management, and environmental protection. Biological Conservation 208: 15-28.
Newman, G., M. Chandler, M. Clyde, B. McGreavy, M. Haklay, H. Ballard, R. Scarpino, R. Hauptfeld, D. Mellor and J. Gallo. 2017. Leveraging the power of place in citizen science for effective conservation decision making. Biological Conservation 208: 55-64.
Perennou, C., N. Sadoul, O. Pineau, A. R. Johnson and H. Hafner. 1996. Management of nest sites for colonial waterbirds. Conservation of Mediterranean wetlands – Number 4. Tour du Valat, Arles, France.
Rabarisoa, R., J. Ramanampamonjy, F. Razafindrajao, L-A. Rene De Roland, F. Jeanne and O. Bacar (in review). Status assessment and population trends of the Madagascar Pond-heron (Ardeola idea) (Hartlaub, 1860), 1993-2016. Waterbirds.
Redman, N., T. Stevenson and J. Fanshawe. 2011. Birds of the Horn of Africa - Ethiopia, Eritrea, Djibouti, Somalia and Socotra. Revised and Expanded Edition. Princeton University Press, Princeton, New Jersey, U.S.A.
Richardson, A. J., I. R. Taylor and J. E. Growns. 2001. The foraging ecology of egrets in rice fields in southern New South Wales, Australia. Waterbirds 24: 255-264.
Rush, S. A., C. Pekarik, D. V. Weseloh, F. Cuthbert, D. Moore and L. Wires. 2015. Changes in heron and egret populations on the Laurentian Great Lakes and connecting channels, 1977-2009. Avian Conservation and Ecology 10(1): 7. [online] Accessed 27 June 2019.
Spies, R. B. and T. Weingartner. 2007. Long-term change. Pages 259-418 in Long-term ecological change in the northern Gulf of Alaska (R. B. Spies, ed.). Elsevier, Amsterdam, The Netherlands.
Stier, A. 2018. Studies of the Agami Heron in French Guiana. In Weseloh, D. V. C. and M. C. Green, (eds.). Research summaries from the Herons of the World Symposium; 2016. Journal of Heron Biology and Conservation 3(5): 10-11. [online] Accessed 3 December 2018.
Sullivan, B. L., T. Phillips, A. A. Dayer, C. L. Wood, A. Farnsworth, M. J. Iliff, I. J. Davies, A. Wiggins, D. Fink, W. M. Hochachka, A. D. Rodewald, K. V. Rosenberg, R. Bonney and S. Kelling. 2017. Using open access observational data for conservation action: A case study for birds. Biological Conservation 208: 5-14.
Tarboton, W. R. 1977. Status of communal herons, ibis and cormorants on the Witwatersrand. South African Journal of Wildlife Research 7: 19-25.
Telfair II, R. C., B. C. Thompson and L. Tschirhart. 2000. Nuisance heronries in Texas: characteristics and management. Second Edition. [online] Accessed 20 September 2018.
Turner, D. 2002. Herons in Africa and the Malagasy region. Pages 123-150 in Heron Conservation (J. A. Kushlan and H. Hafner, eds.). Academic Press, London, U.K.
Turner, D. 2011. Checklist and status of herons in East Africa. Journal of Heron Biology and Conservation 1(9): 1-8. [online] Accessed 20 September 2018.
Tyler, S. J. 2001. A review of waterbird counts in Botswana, 1991-2000. Babbler Special Supplement No. 1. BirdLife Botswana, Gaborone, Botswana.
Underhill, L. G., R. J. M. Crawford, D. M. Harebottle and K. M. C. Tjørve. 2009. The development of the heronry on Robben Island, Western Cape, South Africa, 1980-2005. Pages 217-221 in Final report of BCLME (Benguela Current Large Marine Ecosystem) project on top predators as biological indicators of ecosystem change in the BCLME. (S. Kirkman, ed.). Avian Demography Unit, University of Cape Town, Cape Town, South Africa.
Veen, J., H. Dallmeijer and C. Diagana. 2007. Monitoring colonial nesting birds along the West African Seaboard. Wetlands International Africa, Dakar, Senegal.
Veen, J., W. C. Mullié, C. M. Sylla, P. Robinson and M. S. Diop. 2011. Suivi de la reproduction 2011 des colonies d’oiseaux marins de l’Ile aux Oiseaux, Parc National du Delta du Saloum. Rapport interne soumis à la Fondation Internationale du Banc d’Arguin (FIBA) Dakar/Wenum Wiesel, August 2011. Dakar, Senegal.
Wetlands International. 2012. Waterbird population estimates, fifth edition. Summary report. Wetlands International, Wageningen, The Netherlands.
Whittington-Jones, C. (ed.). 2014. Humans and birds – A guide to problem resolution and mitigation. Endangered Wildlife Trust, Johannesburg, South Africa.
Yanosky, A. 2018. Heron conservation in Paraguay. In Weseloh, D. V. C. and M. C. Green, (eds.). Research summaries from the Herons of the World Symposium; 2016. Journal of Heron Biology and Conservation 3(5): 11-12. [online] Accessed 3 December 2018.